|
|
|
|
|
|
|
|
 |
Esta glândula
pineal normal foi obtida da autópsia de um paciente do sexo masculino,
38 anos, portador de síndrome hemofagocítica. O cérebro
era macro- e microscopicamente normal.
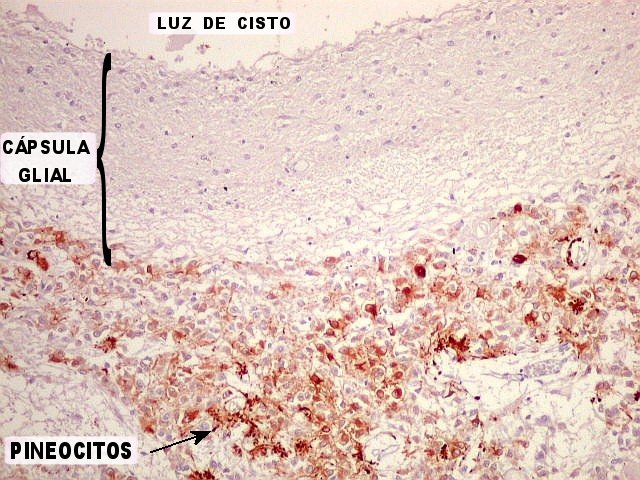
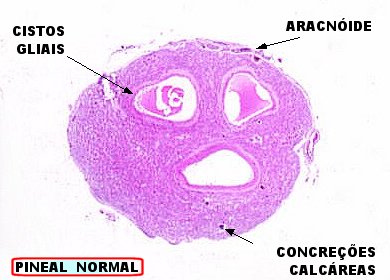
A imagem escaneada da lâmina mostra três cistos de parede glial no parênquima da glândula, dois contendo material róseo amorfo. Externamente, a glândula é revestida por fina camada de aracnóide. Algumas pequenas calcificações (corpora arenacea) são visíveis. |
 |
|
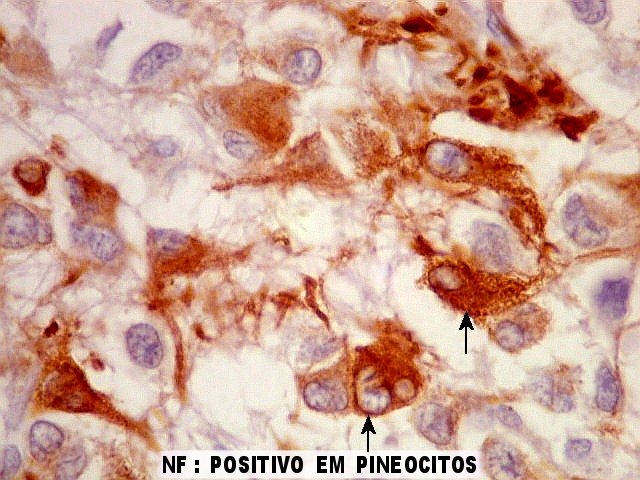
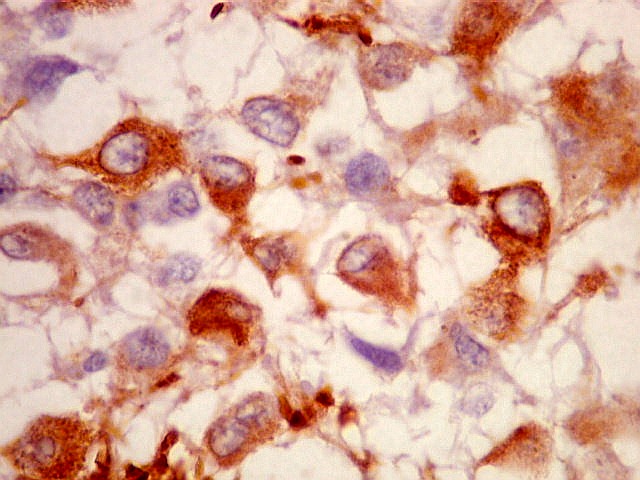
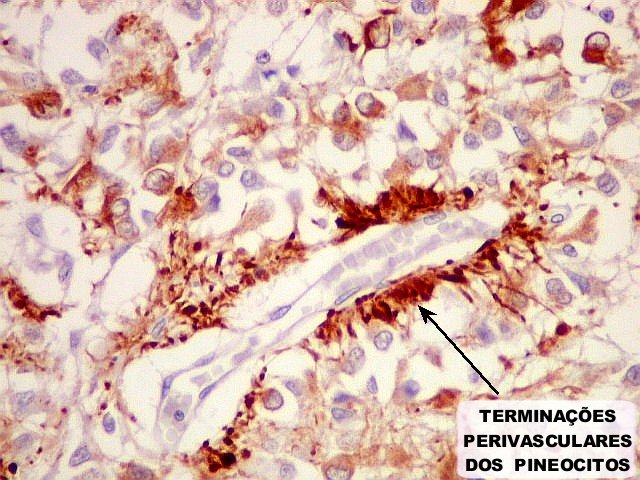
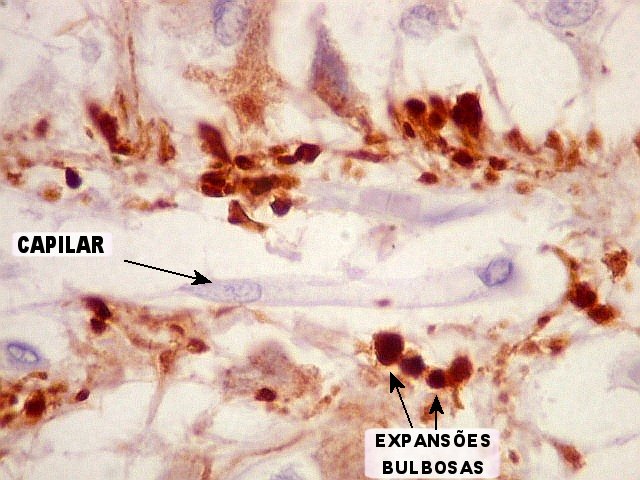
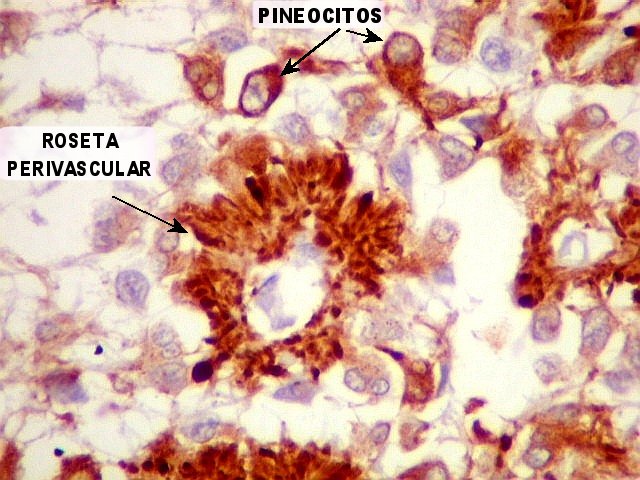
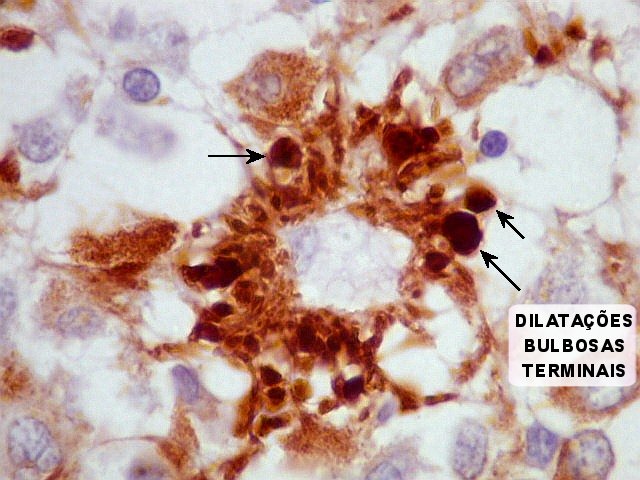
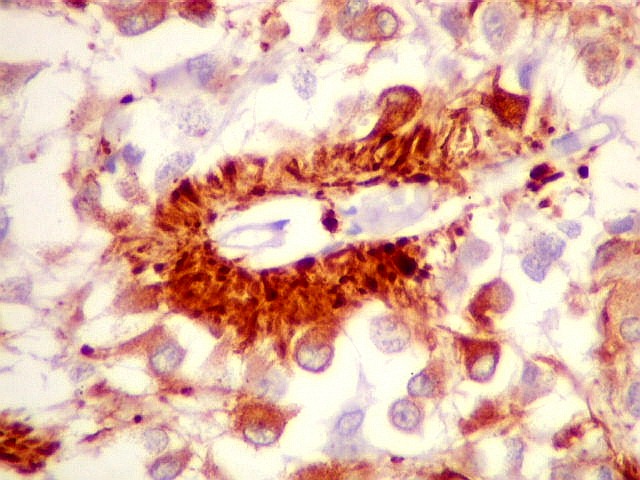
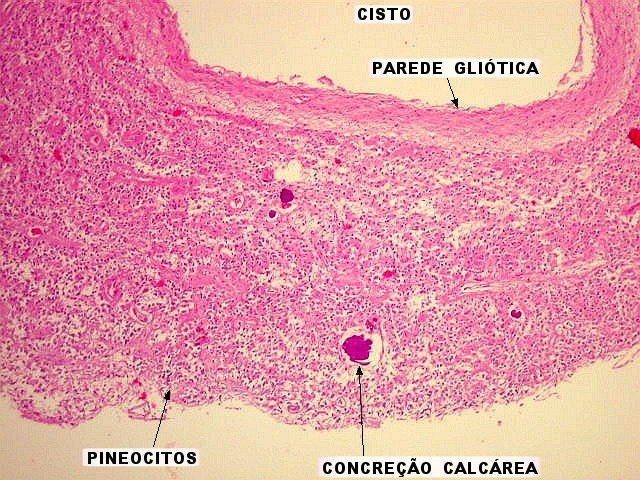
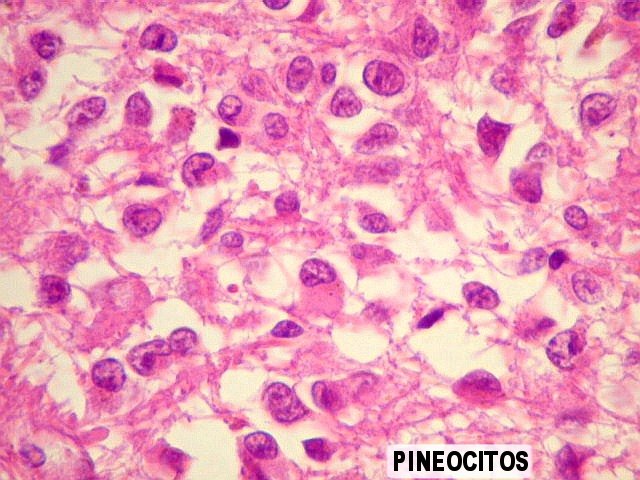
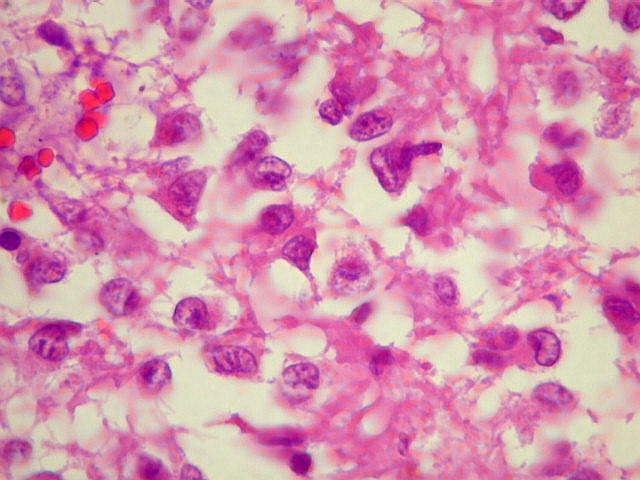
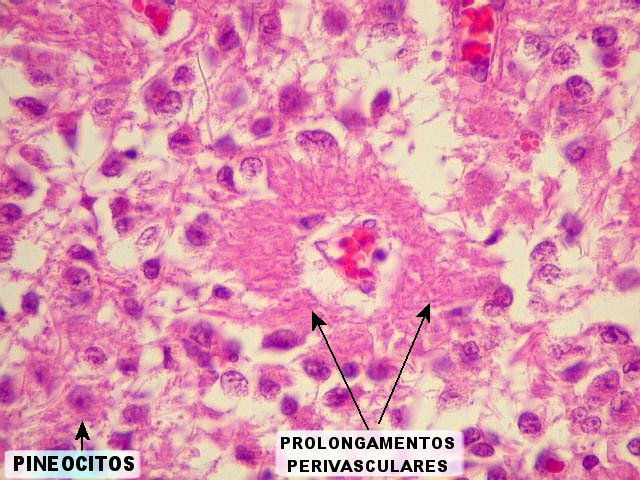
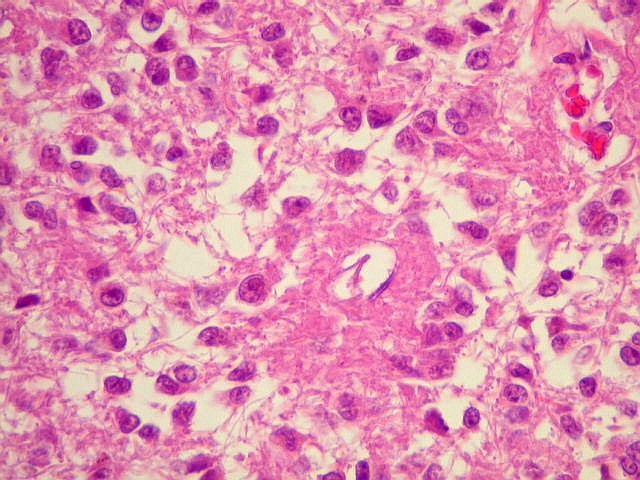
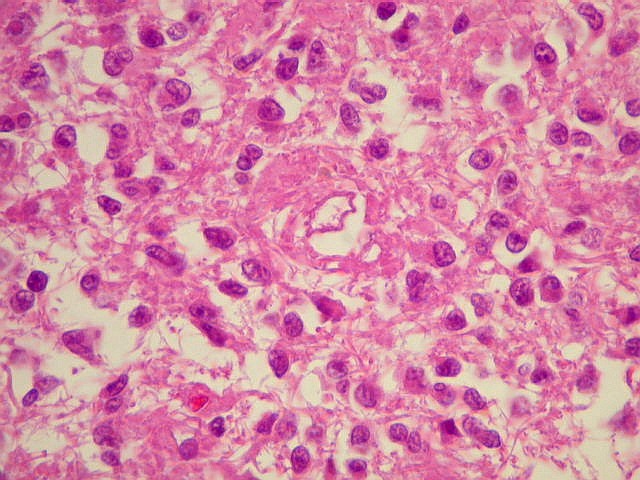
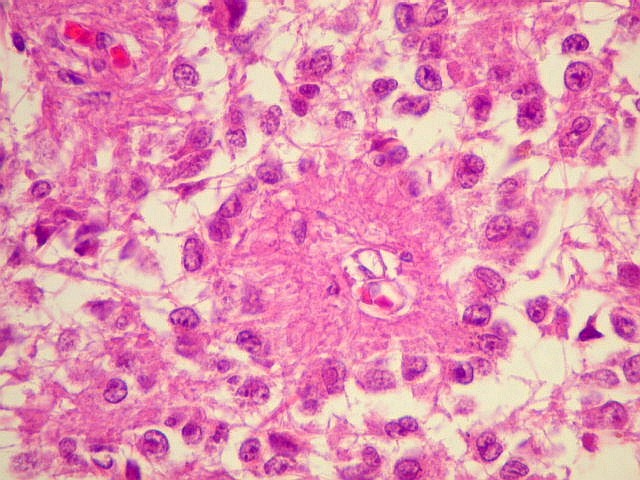
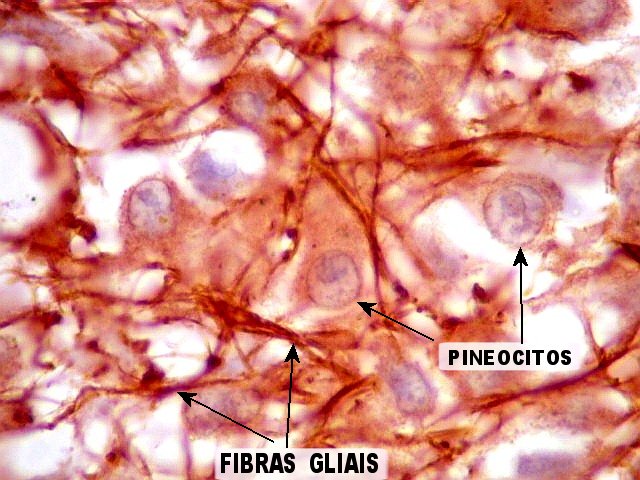
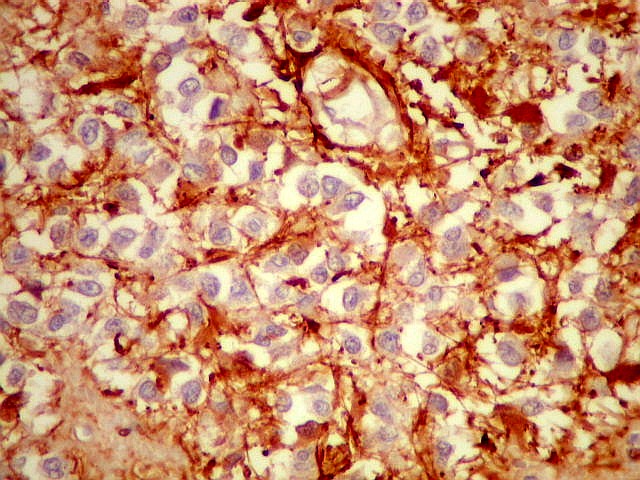
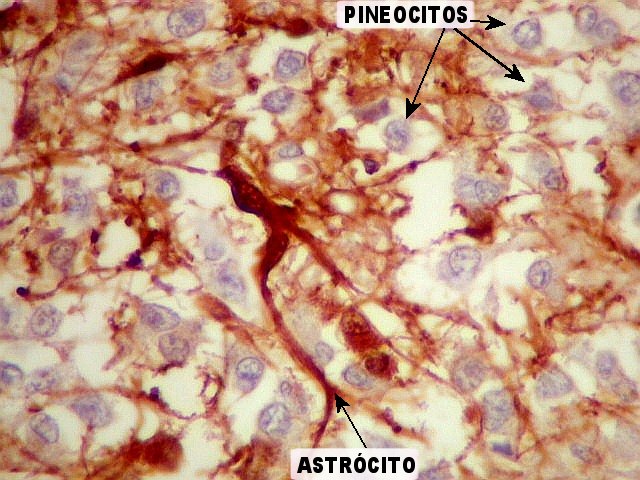
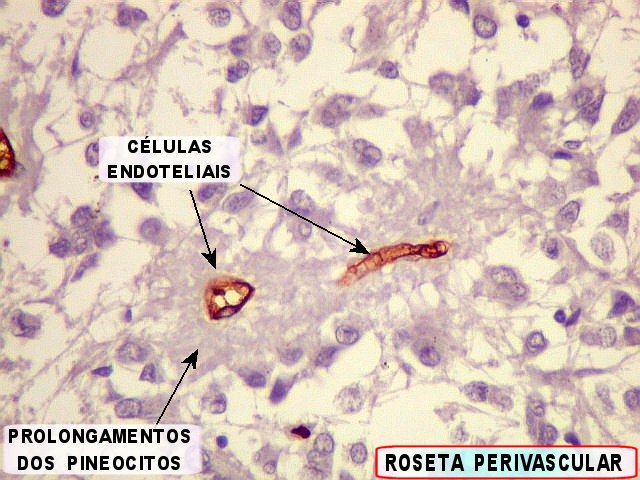
| Pineocitos. São as células principais da pineal. São derivados de neurônios, mas não têm axônio. Têm formato mais ou menos poligonal, com prolongamentos. Alguns destes terminam na parede de pequenos vasos, com expansões terminais em forma de bulbo, demonstráveis por imunohistoquímica para marcadores neuronais, como NF, SNF e CGR. | |
 |
 |
 |
 |
 |
 |
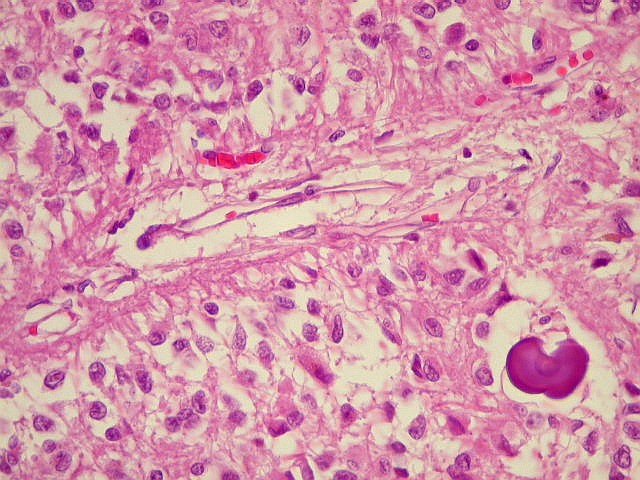
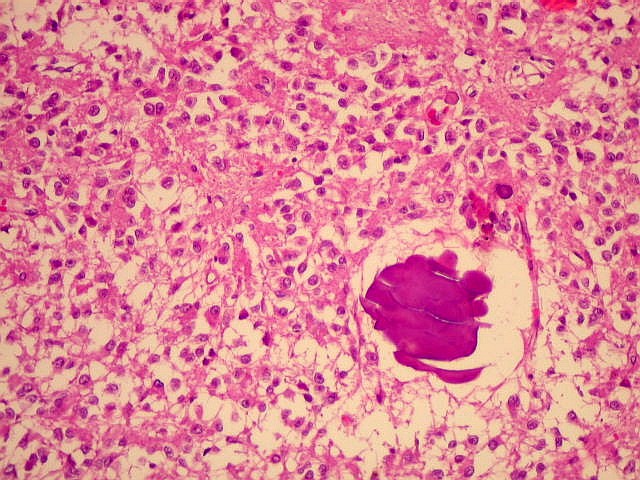
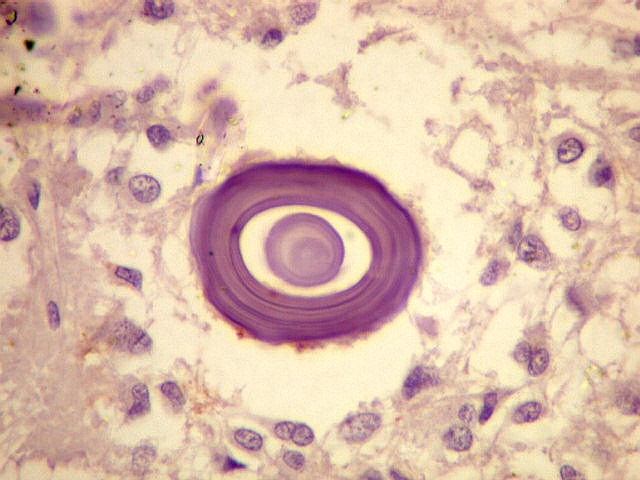
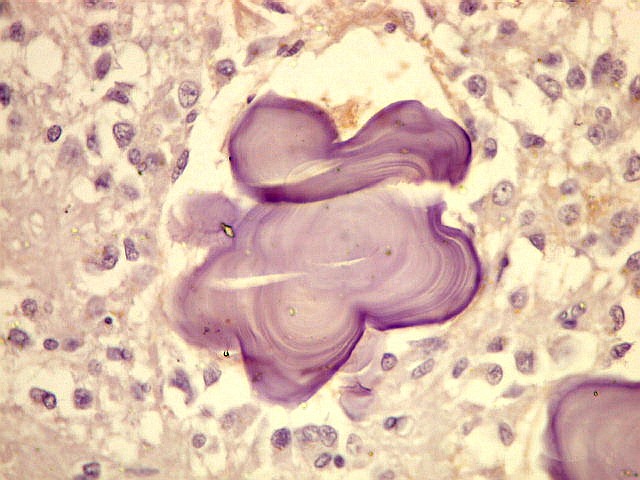
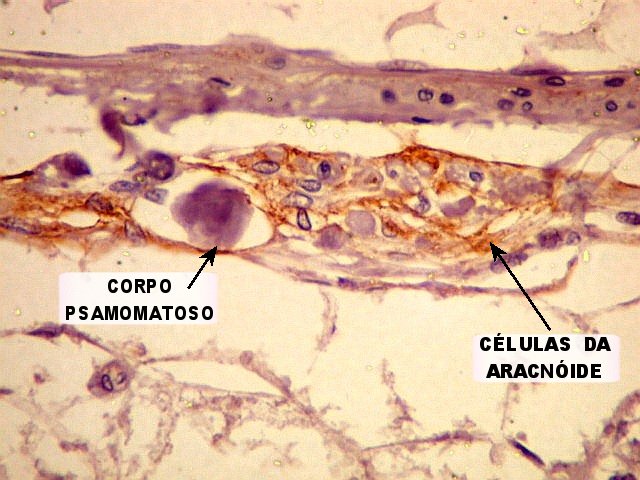
| Concreções calcáreas ou corpora arenacea aumentam em número com a idade. Fileira de cima, HE; fileira de baixo - imunohistoquímica para EMA (negativa no parênquima pineal, mas a contracoloração pela hematoxilina demonstra bem as concreções). | |
 |
 |
 |
 |
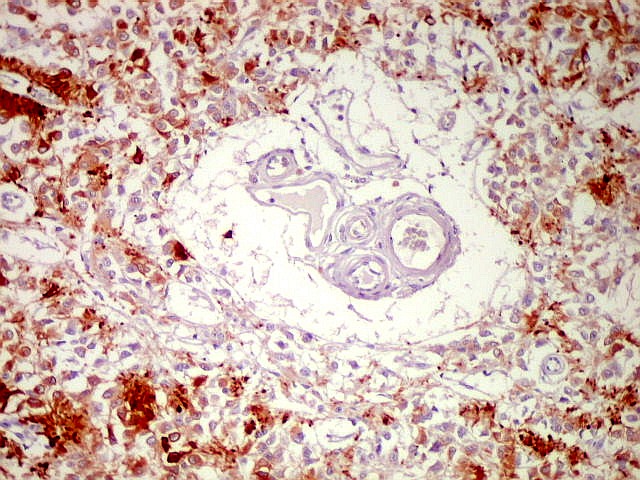
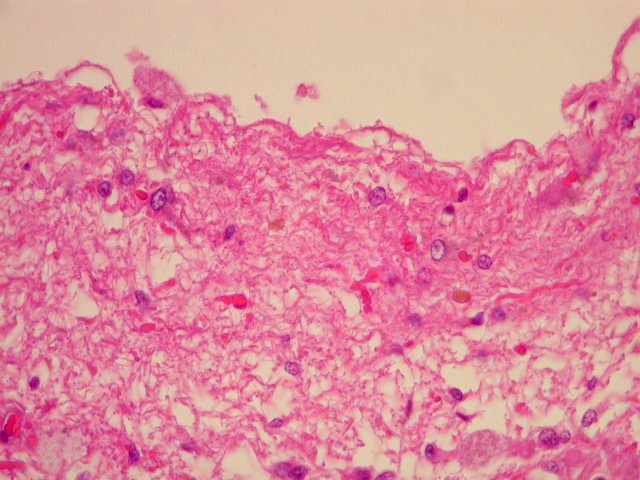
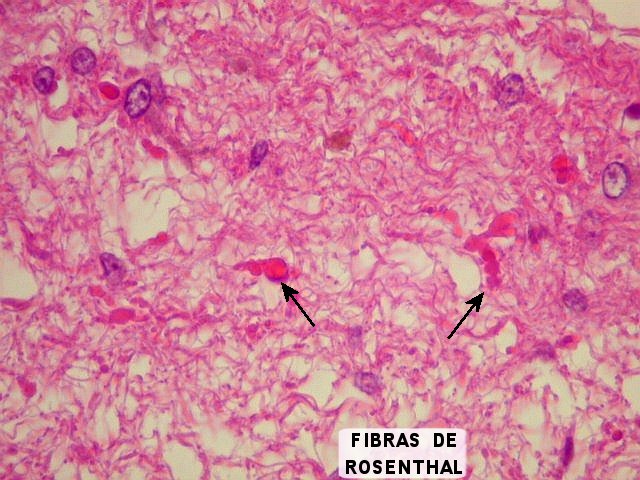
| Cistos. Sua parede é densamente gliótica, constituída por astrócitos com prolongamentos ondulados e tortuosos, lembrando um astrocitoma pilocítico. Há fibras de Rosenthal. | |
 |
 |
..
|
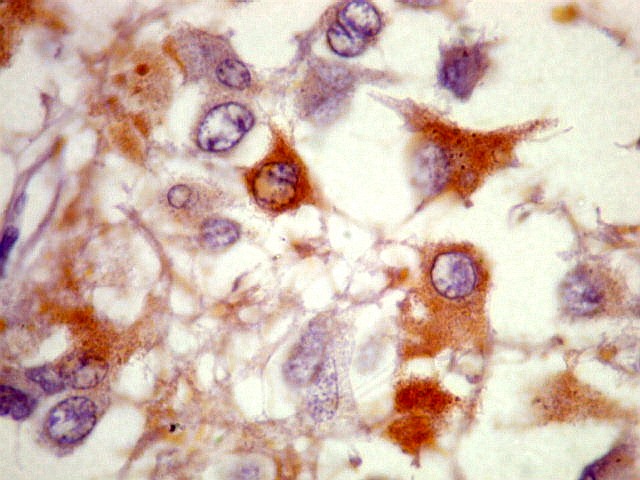
(marcadores neuronais): NSE, NF, SNF, CGR. |
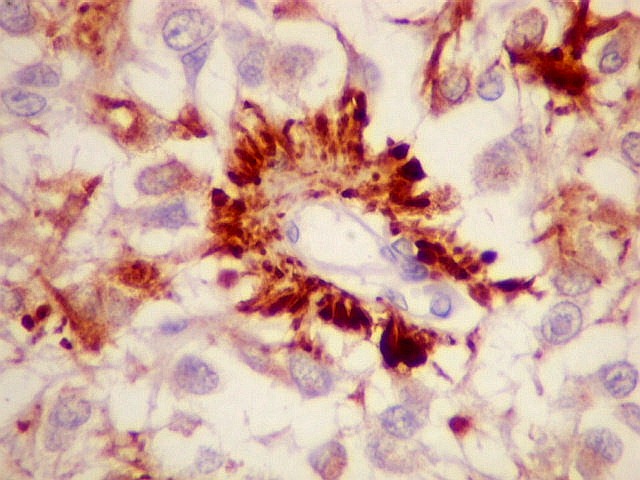
| NSE. Positivo nos pineocitos, negativo na margem glial dos cistos pineais. | |
 |
 |
 |
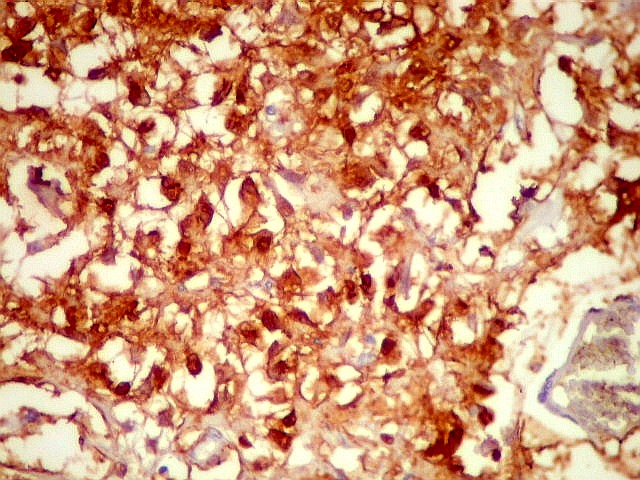 |
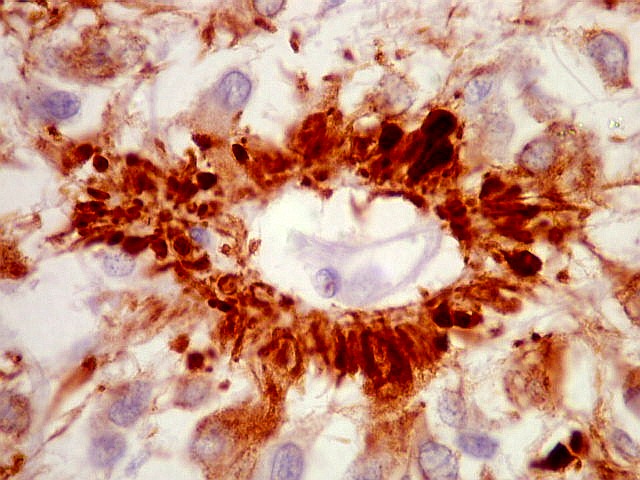
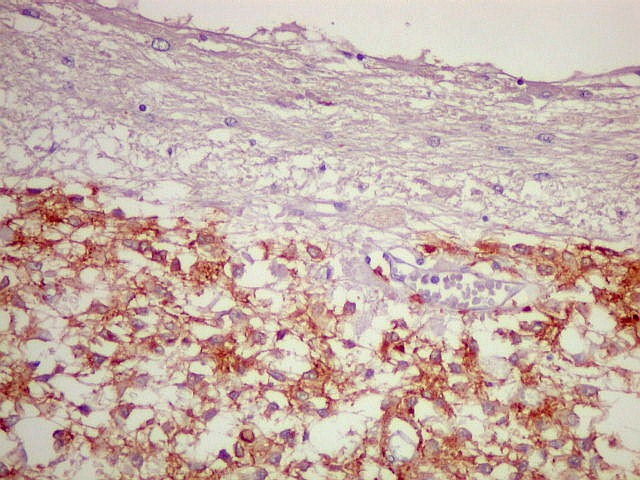
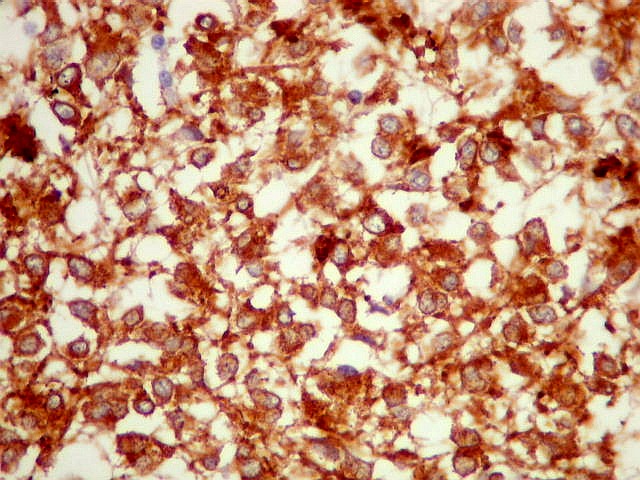
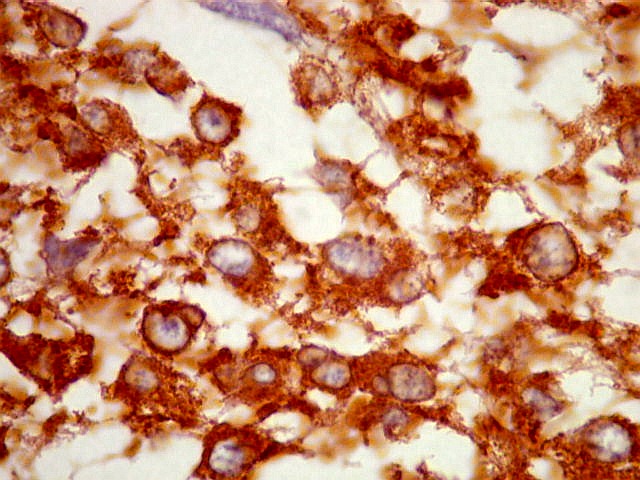
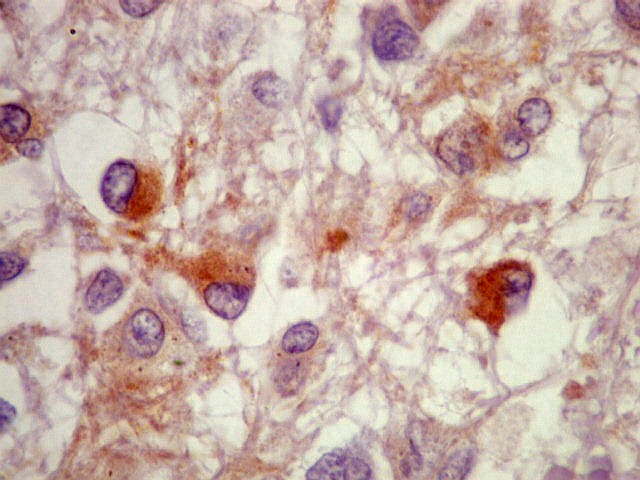
| SNF. Resultados análogos aos com NF, acima. | |
 |
 |
 |
 |
 |
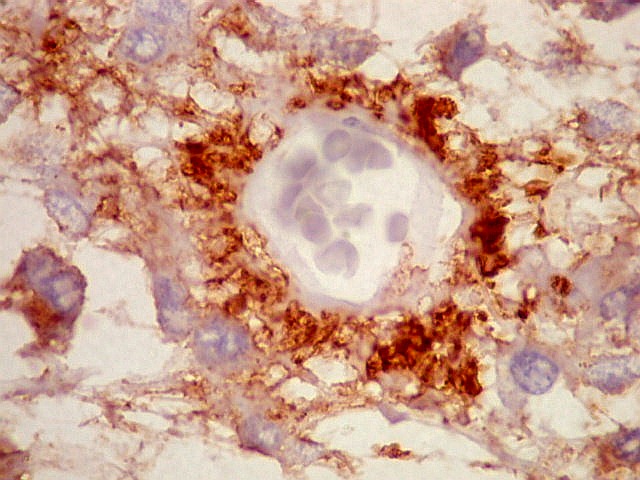 |
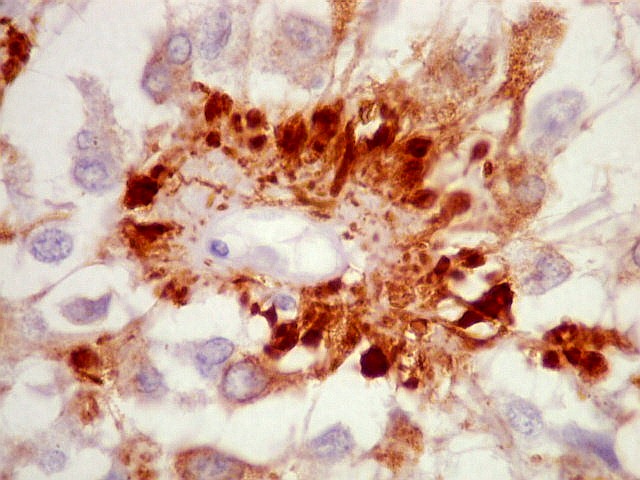
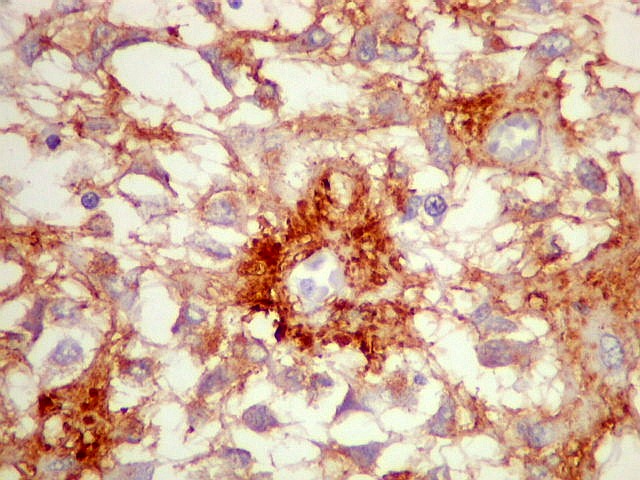
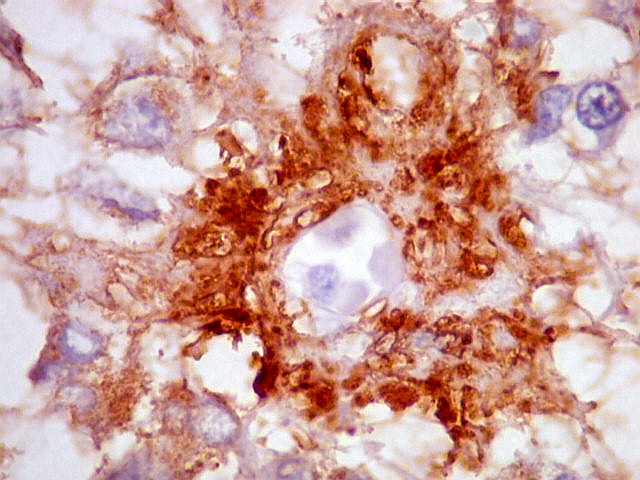
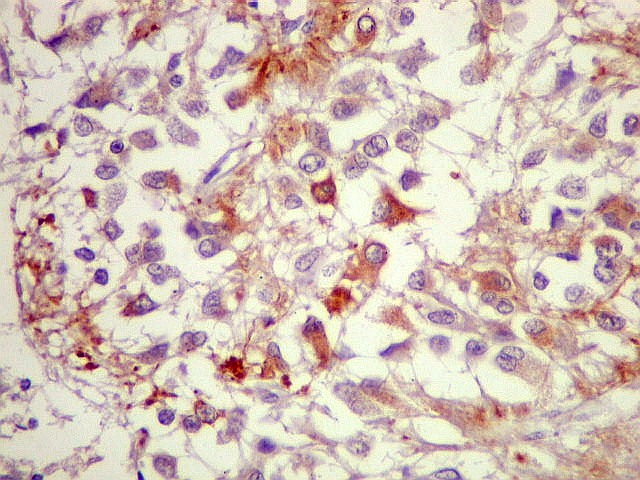
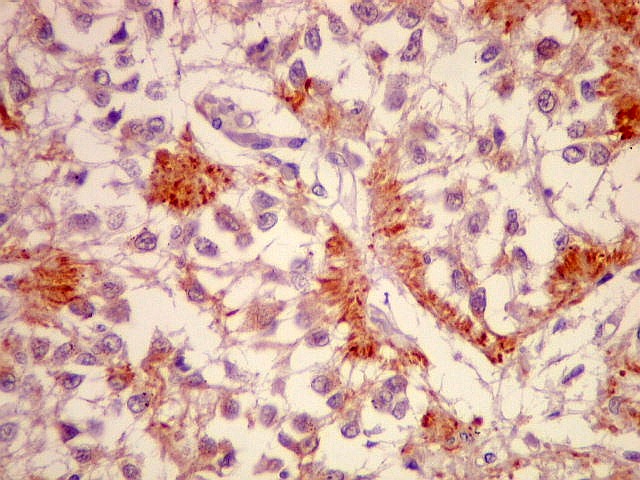
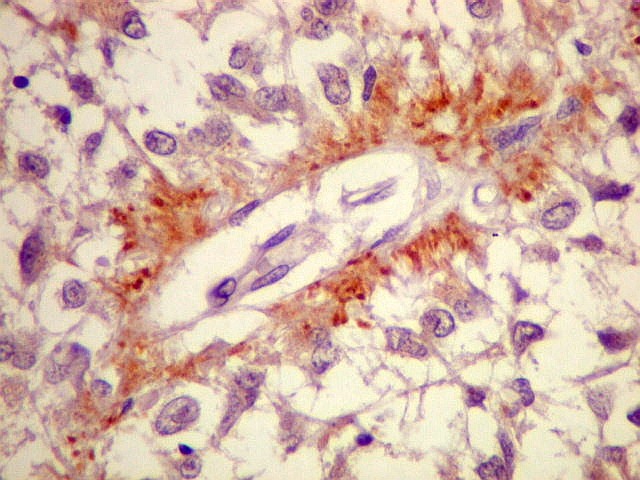
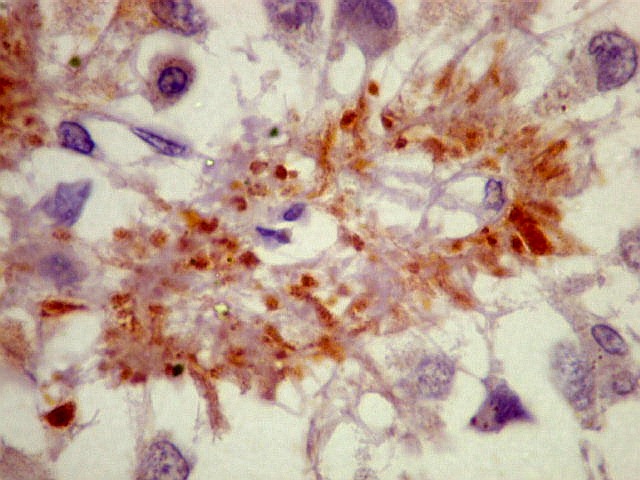
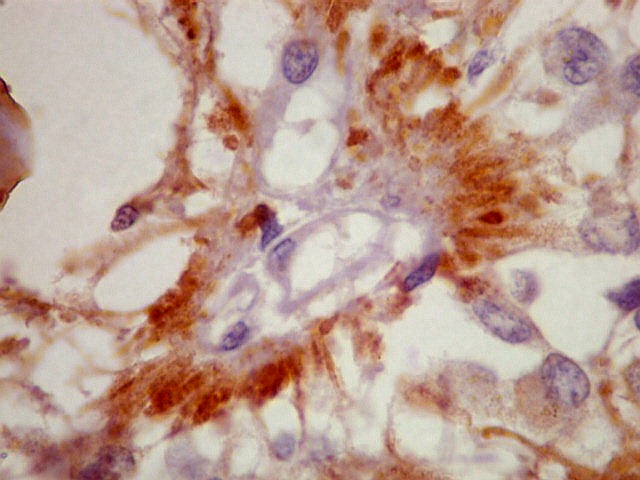
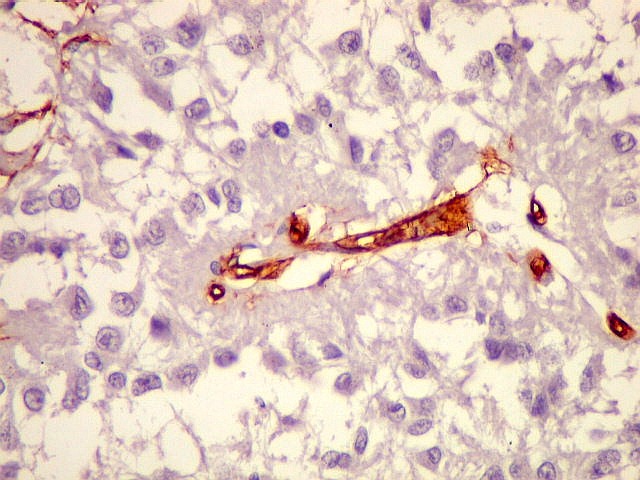
| CGR. Demonstra pineocitos e rosetas perivasculares, como NF e SNF, acima. | |
 |
|
 |
 |
 |
 |
 |
 |
..
|
vasos (VIM, CD34) e meninge (EMA). |
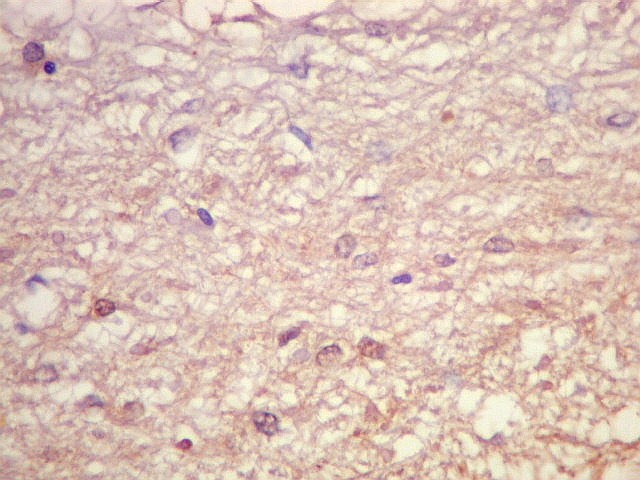
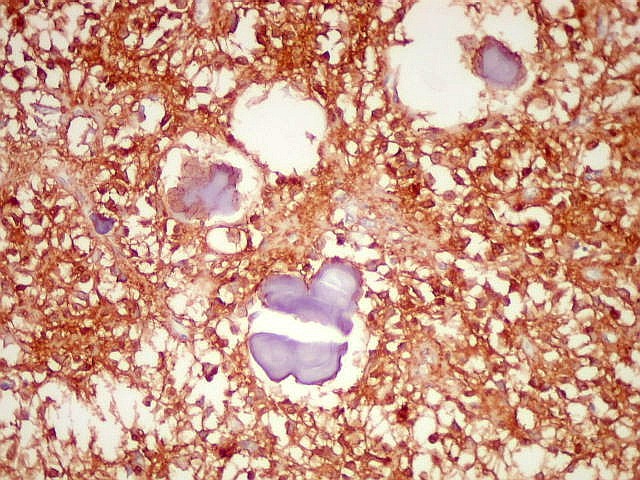
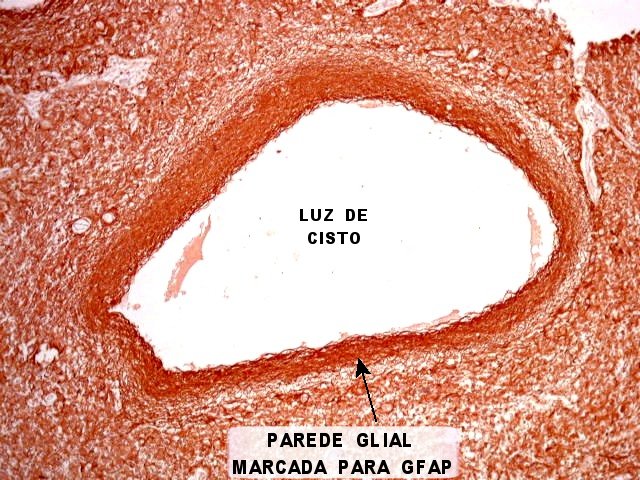
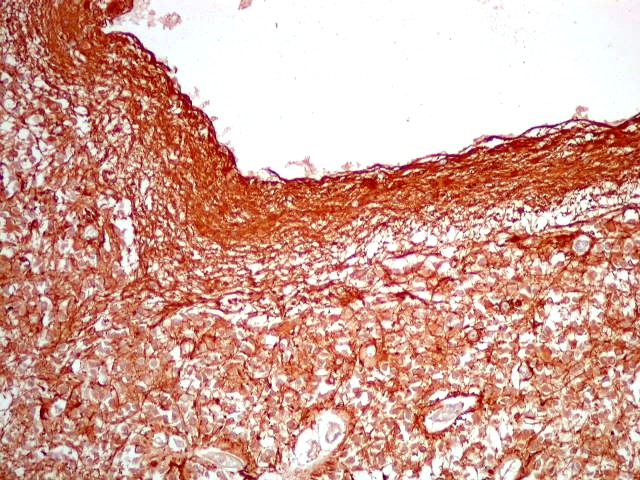
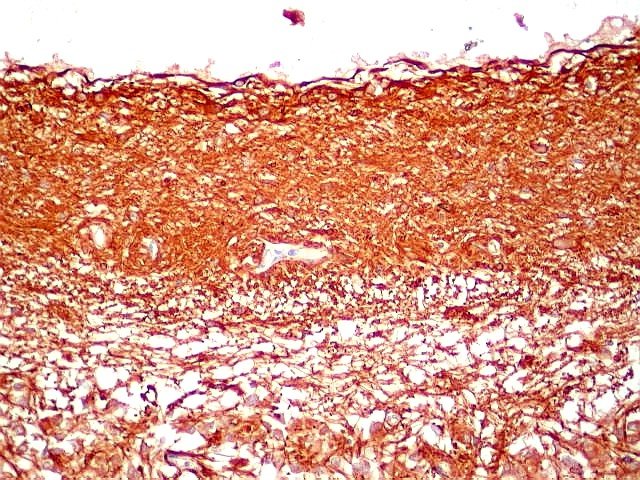
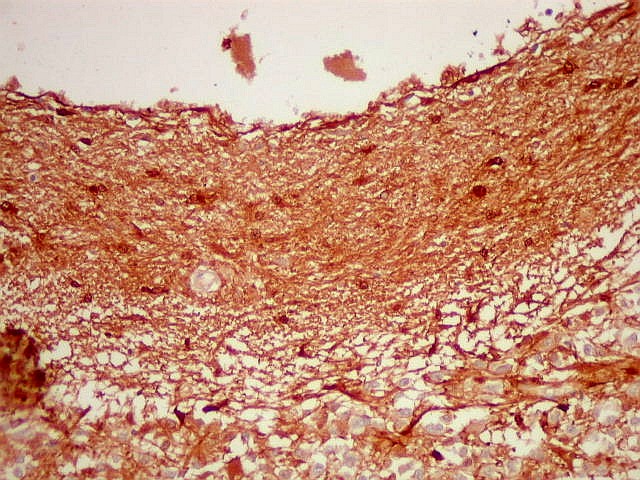
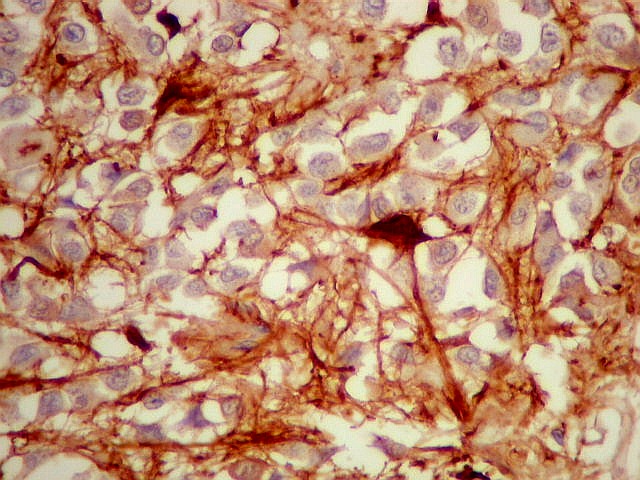
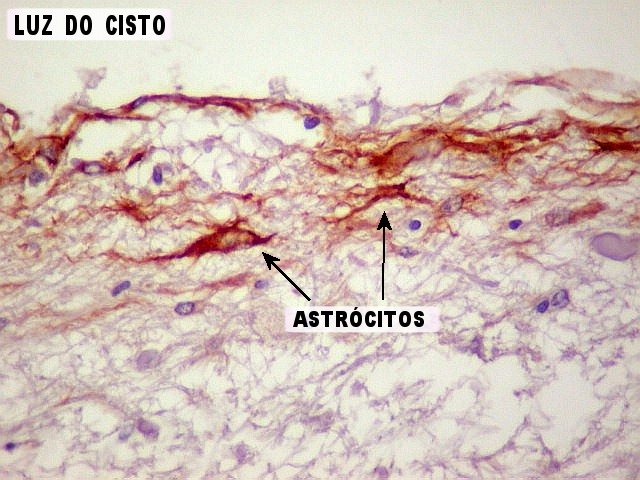
| GFAP. Marca a parede glial dos cistos e os astrócitos que dão suporte aos pineocitos. Os astrócitos constituem 5-10% do total de células da pineal. | |
 |
 |
 |
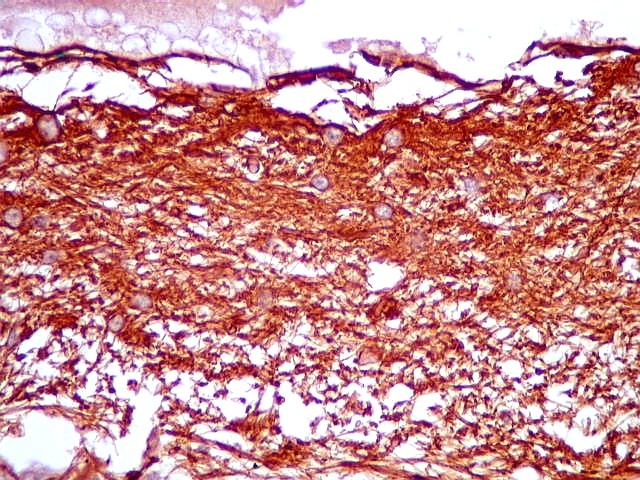 |
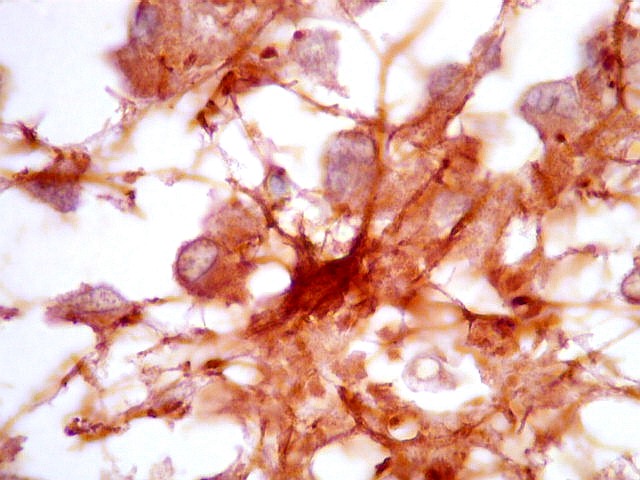
| Astrócitos. Destacam-se dos pineocitos por seus prolongamentos longos e marcação mais forte. Os pineocitos neste exemplo mostram reação de fundo (inespecífica). | |
 |
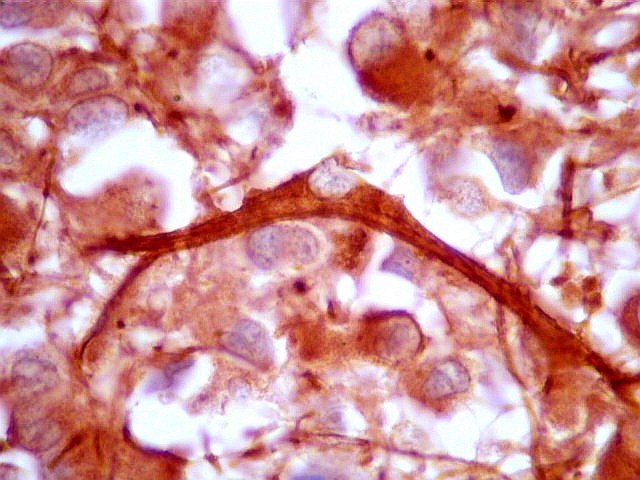 |
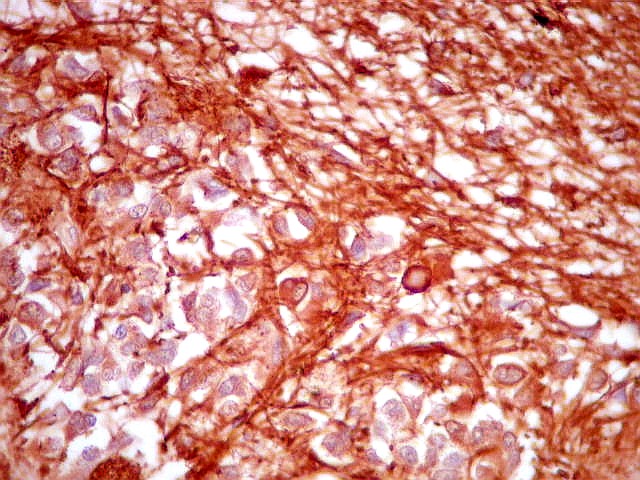
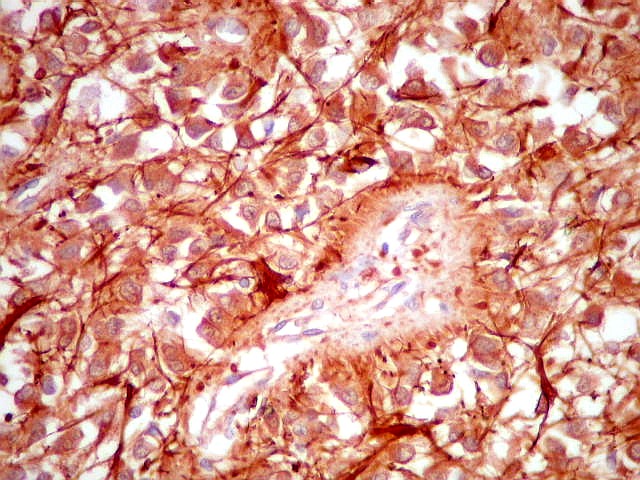
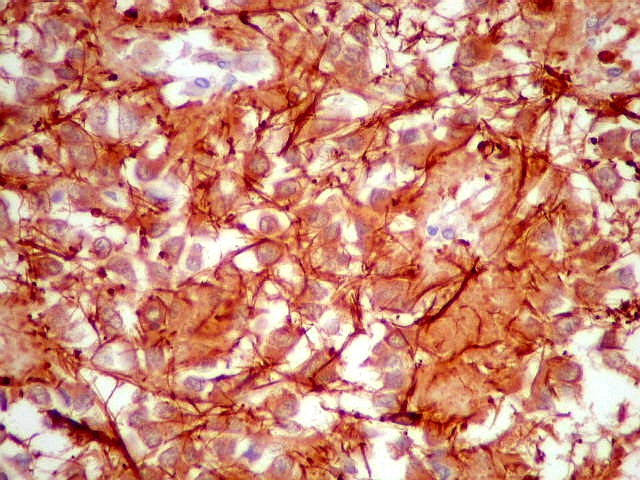
| Fibras gliais. Os prolongamentos astrocitários formam delicada trama entre os pineocitos. | |
 |
 |
 |
 |
| S100. Também demonstra o componente astrocitário da pineal (resultado semelhante ao com GFAP). | |
 |
 |
 |
 |
| VIM. Positiva em astrócitos fibrosos da parede dos cistos, e em células endoteliais. Os astrócitos do parênquima não são marcados. | |
 |
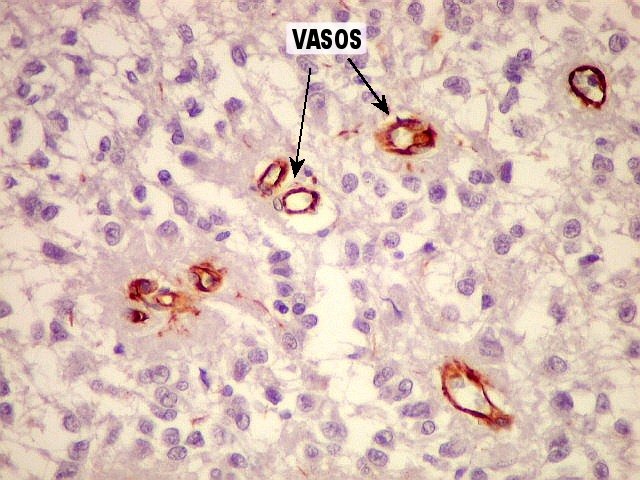 |
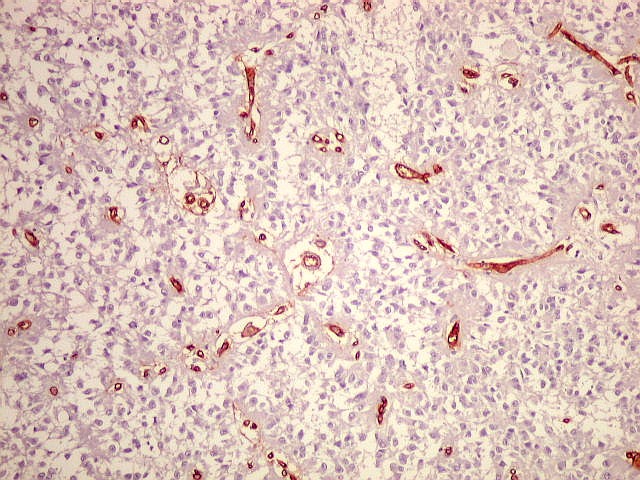
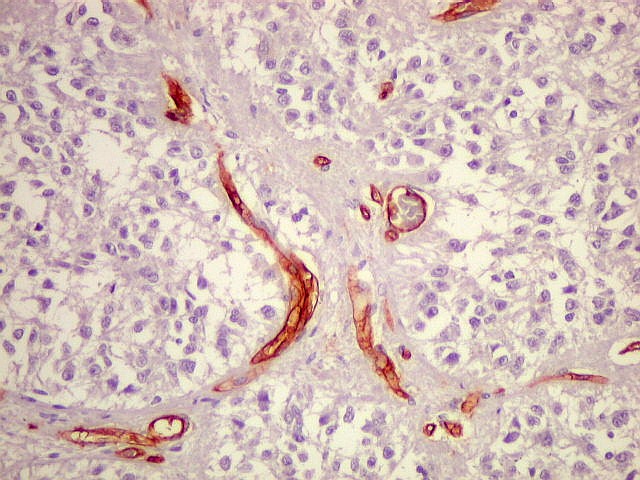
| CD34. Positivo em células endoteliais, demonstra o arcabouço vascular. Os pineocitos são negativos. | |
 |
 |
 |
 |
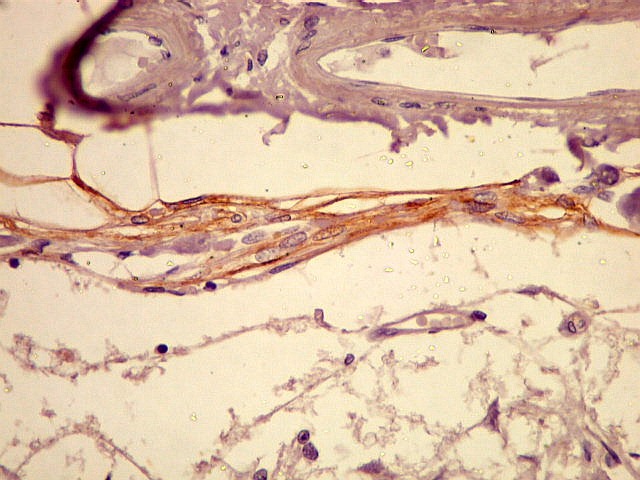
| EMA. Positivo nas células da aracnóide que circundam a glândula pineal. | |
 |
 |
| Ciclo circadiano.
A vida na terra desenvolveu-se na vigência de ciclos de dia e noite e a maioria dos organismos tem um ritmo interno que dita diferentes comportamentos de acordo com a hora do dia. Estes comportamentos incluem o ritmo sono-vigília, além de alterações cíclicas em atividades enzimáticas, liberação de hormônios e temperatura corporal. Tais ritmos diurnos são controlados por osciladores internos chamados relógios circadianos (circadian clocks). Esses relógios internos têm um ciclo intrínseco de mais ou menos 24 horas, um pouco mais ou um pouco menos, que continua a existir mesmo na ausência de influências externas. Contudo, como um relógio biológico não pode ser perfeitamente acurado, precisa ser resetado ou acertado com base em indícios ambientais, como a luz do dia. Estes estímulos externos mantêm o organismo em sincronia com seu ambiente. No caso de mudanças bruscas, como de fuso horário, o relógio interno é gradualmente ajustado, às vezes à custa de certo desconforto (jet lag). O mecanismo de oscilação biológico ocorre individualmente em células do núcleo supraquiasmático no hipotálamo. Se estas células forem removidas do corpo e mantidas em cultura, continuam a mostrar um padrão cíclico de expressão de certos genes, que dura cerca de 24 horas. No indivíduo normal, estas células (chamadas SCN para suprachiasmatic nucleus) recebem sinalização das variações de luminosidade a partir da retina, e passam informações a outros tecidos, como a glândula pineal, que, por sua vez, sinaliza ainda outros tecidos através da liberação do hormônio melatonina. O mecanismo molecular do relógio biológico ainda é pouco compreendido. Em Drosophila, deve-se a variações da expressão de certos genes, que são controlados através de alças de retroalimentação negativas com retardo. Quando os genes são transcritos, os seus produtos acumulam, migram para o núcleo onde inibem a transcrição dos mesmos genes. A parada da transcrição e a degradação dos produtos libera novamente a transcrição e assim por diante. |
 |
| PINEAL
NORMAL
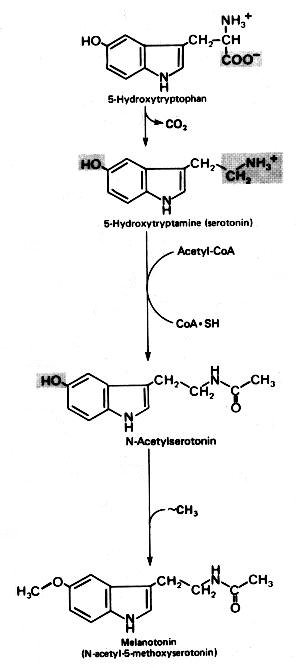
A glândula pineal (sinônimos: epífise, conário) começa a desenvolver-se no segundo mês de vida intrauterina como uma pequena invaginação no teto do diencéfalo. Na vida fetal e na infância precoce tem estrutura lobulada, que fica menos nítida no adulto. As células parenquimatosas da pineal são chamadas pinealócitos ou pineocitos. São uma forma especializada de neurônios relacionados aos fotoreceptores retinianos. Não têm axônios, mas contêm no citoplasma numerosos grânulos secretórios e prolongamentos citoplasmáticos de comprimento variado. Os mais longos dirigem-se a vasos, onde terminam em dilatações bulbosas (clubbed endings). Esses processos celulares, que são demonstrados por técnicas argênticas como o carbonato de prata amoniacal de Del Rio Hortega, são escassos antes da idade de 8 anos. Atualmente, imunohistoquímica para antígenos neuronais, como neurofilamento, sinaptofisina e cromogranina, também pode ser usada. Em roedores, na fase fetal e em recém nascidos, os pineocitos têm características ultraestruturais que lembram células fotoreceptoras, recordando a filogenia da pineal, que, em espécies inferiores, é um órgão fotoreceptor (o terceiro olho). Após o nascimento estas feições desaparecem. No homem, não foi encontrada expressão transitória de características fotoreceptoras na pineal. Contudo, em mamíferos, os pineocitos compartilham com as células retinianas a capacidade para expressar o chamado antígeno S, uma proteína de 48 kD que se liga à rodopsina, e que pode estar envolvida nos eventos iniciais da transdução da luz em sinais químicos. O antígeno S pode ser demonstrado por imunohistoquímica em um pequeno número de pineocitos na pineal humana fetal, infantil e adulta. Também expressam a enzima rodopsina quinase, outra substância encontrada em células fotoreceptoras bem diferenciadas. Além dos pineocitos, a pineal contém astrócitos (cerca de 5 a 10% das células), que parcialmente circundam e separam os pineocitos. O pedúnculo da pineal é constituído em parte por prolongamentos destas células. Com imunohistoquímica, os astrócitos de suporte são revelados por GFAP e proteína S100. Os capilares da pineal são forrados por células endoteliais finas e às vezes fenestradas, apoiadas sobre lâmina basal fina e às vezes incompleta. A inervação da pineal é feita por axônios amielínicos simpáticos, noradrenérgicos, que contêm vesículas de centro denso em suas expansões terminais. Essas fibras originam-se no gânglio cervical superior, e penetram na pineal através dos chamados nervi conarii. Estes penetram pela porção dorsal da glândula vindos da tenda do cerebelo, onde correm em posição subendotelial na parede do seio reto. Uma feição característica da pineal, que começa a aparecer na infância e aumenta com a idade, são microcalcificações, chamadas de corpora arenacea, acervuli cerebri ou areia cerebral. Têm estrutura lamelar concêntrica e parecem ser formados por complexos de íons cálcio com proteínas, associados à atividade secretória (por exocitose) dos pineocitos. As calcificações podem ser visíveis já em radiografias simples do crânio, e na era pré tomografia computadorizada eram um marco importante da linha média cerebral. Também com a idade tende a haver gliose e aparecimento de cistos. Estes têm paredes glióticas que podem conter fibras de Rosenthal (ver acima). Fisiologia da pineal. Há evidência de que a pineal de mamíferos é uma glândula endócrina que modula a atividade de outras glândulas, como a adenohipófise, neurohipófise, pâncreas endócrino, paratiróides, córtex e medula da adrenal e gônadas (portanto, um transdutor neuroendócrino). De modo geral, os efeitos das secreções da pineal são inibitórios. A pineal secreta melatonina (uma indolamina, ver fórmula acima) e hormônios polipeptídicos, que parecem diminuir a síntese e liberação de vários hormônios na adenohipófise, seja por ação direta sobre as células secretoras, seja inibindo produção de fatores liberadores no hipotálamo. As secreções da pineal atingem seus alvos através do líquor ou por via hematogênica. Nas gônadas, a melatonina parece inibir a atividade esteroidogênica. Tanto a melatonina como as enzimas necessárias para sua síntese, como a serotonina N-acetil transferase, mostram variação circadiana de concentração e atividade na pineal de vários animais. No rato, há um oscilador endógeno no núcleo supraquiasmático do hipotálamo, cuja ritmicidade intrínseca seria responsável pelo comportamento cíclico da pineal. Os neurônios do núcleo supraquiasmático são supridos diretamente por axônios de células retinianas, informando-os sobre alterações no nível de luminosidade ambiente. No rato, a ligação do núcleo supraquiasmático com a pineal passa pelos núcleos reticulares do tegmento (tronco cerebral), que mandam fibras para a coluna intermédio-lateral da medula torácica alta. Daí, fibras simpáticas pré-ganglionares vão ao gânglio cervical superior, e deste, fibras pós-ganglionares vão até a pineal passando pela tenda do cerebelo e nervos conários (ver acima) e formam terminações bulbosas no corpo dos pineocitos. A liberação de catecolaminas destas fibras causa um aumento na produção de AMP-cíclico nos pineocitos que, por sua vez, causa um aumento de 70 a 100 vezes na atividade da enzima serotonina N-acetil transferase, e é responsável pelas alterações diárias da produção de melatonina e de seus níveis no sangue. O nível aumenta durante a noite até 10 vezes o nível basal e cai durante o dia. Em animais que apresentam alterações de comportamento sazonais (de acordo com a estação do ano), a pineal parece responder a variações no comprimento dos dias e das noites. É importante lembrar que estes mecanismos foram estudados principalmente no rato e ainda não estão totalmente estabelecidos no homem. Fontes: Alberts B et al. Molecular Biology of the Cell. 4th Ed. Garland Science, New York, 2002. pp. 421. Fuller GN, Burger PC. Central Nervous System. in Sternberg SS (ed) Histology for Pathologists. 2nd Ed. Lippincott-Raven, Philadelphia, 1997. pp 268-70. Murray RK et al. (eds) Harper's Biochemistry, 23rd Ed. Appleton Lange, Norwalk CT., 1993. p. 332. Russell DS, Rubinstein LJ. Pathology of Tumours of the Nervous System. 5th Ed. Arnold, London, 1989. p.380-2. Williams PL, Warwick R (eds). Grays Anatomy, 36th Ed. Churchill Livingstone, London, 1980. p. 1445-8. |
| Tumores germinativos_: | Neuroimagem | Neuropatologia | Tumores do parênquima pineal : | Neuroimagem | Neuropatologia |
| Banco de imagens : | Pineal normal,
HE e imunohistoquímica |
Tumores da pineal | Textos ilustrados linkados: | Tumores germinativos do SNC | Tumores do parênquima pineal |
|
|
| Módulo Neuro - Página Inicial | Outros módulos | e-mails : gradanat@fcm.unicamp.br___gradanat@unicamp.br |