|
|
|
|
|
|
|
|
| Série de 11 páginas sobre histologia e imunohistoquímica do hipocampo humano normal em material de autópsia.Página índice da série. Clique para HE, LFB-Nissl, imunohistoquímica para neurônios : MAP2, NF, SNF, cromogranina; para astrócitos : GFAP, VIM; para vasos : CD34; para micróglia : CD68, HAM56. Texto sobre estrutura do hipocampo, texto e esquema sobre conexões do hipocampo. |
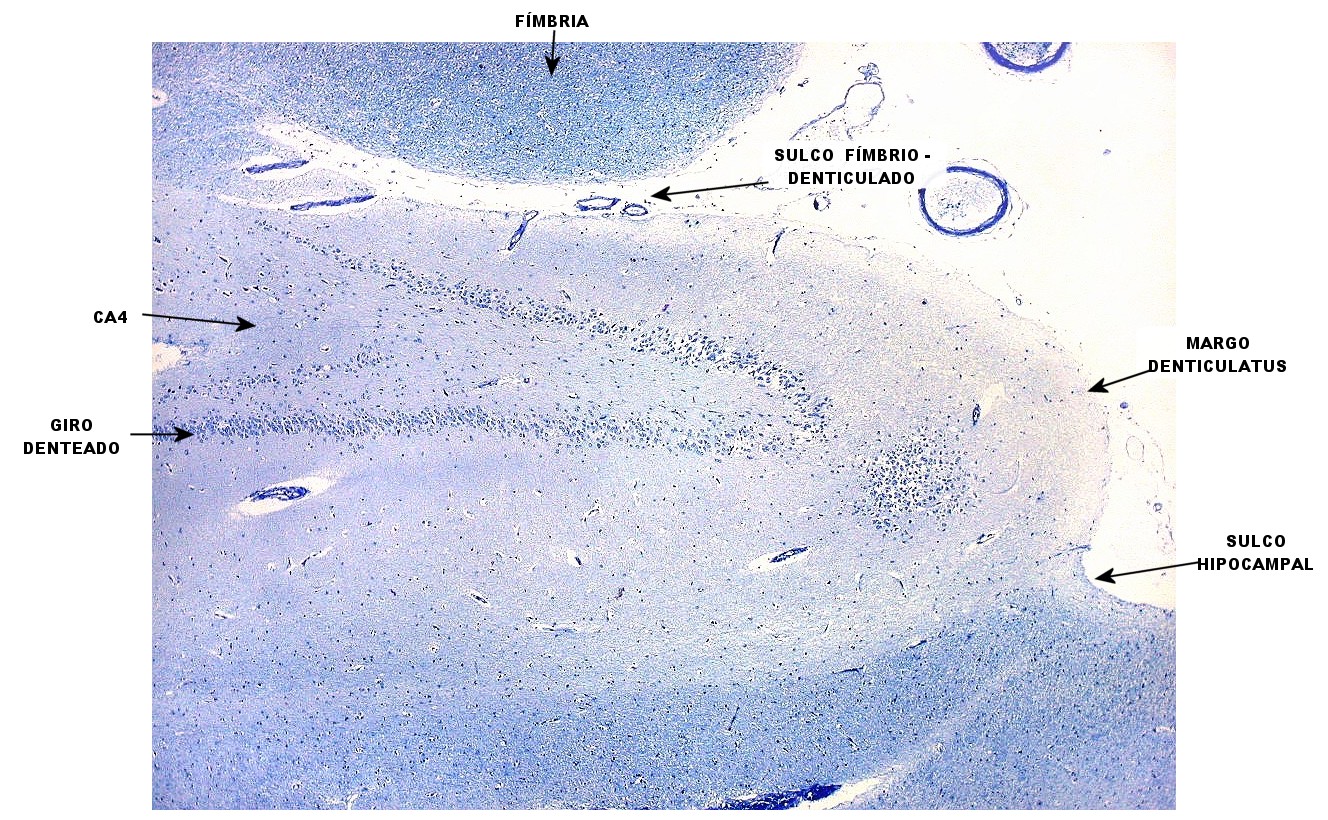
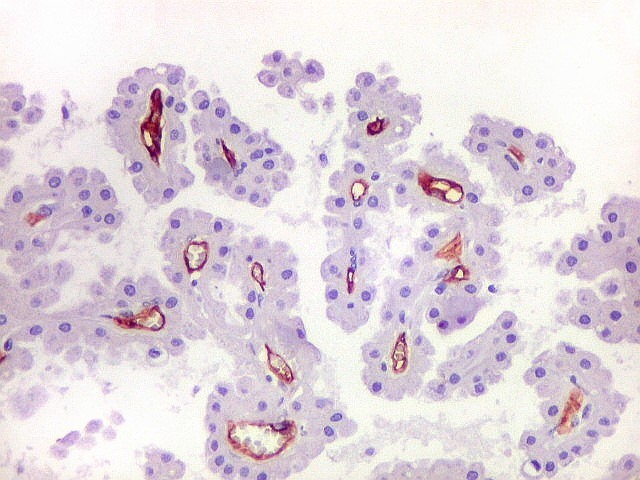
| Destaques do LFB - Nissl. | ||
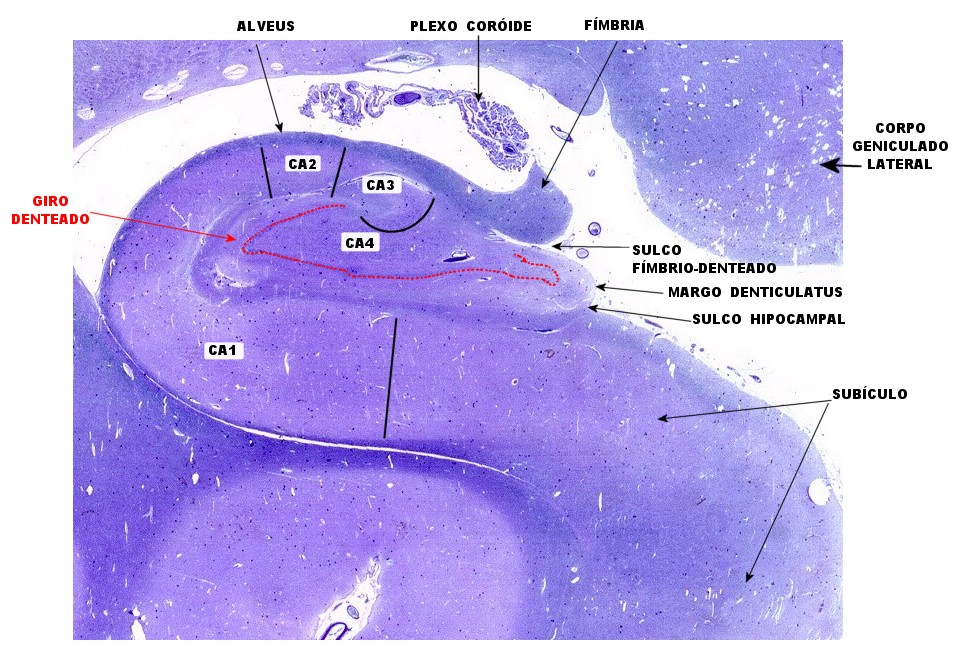
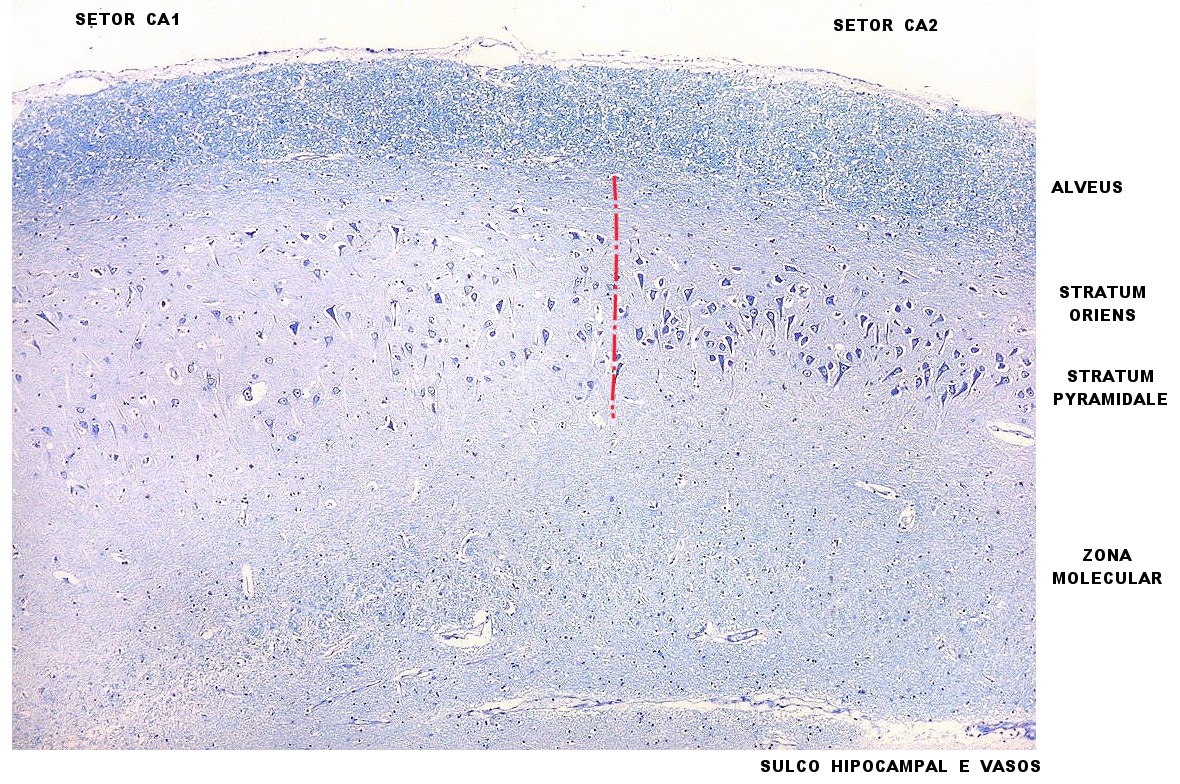
| Lâmina escaneada. | Setores do hipocampo : CA1. CA1 alto, transição CA1-CA2. | CA2 |
 |
 |
 |
| CA3 | CA4 | Giro denteado |
 |
 |
 |
| Subículo | Clusters de neurônios do giro parahipocampal. Fibras mielínicas do subículo | Sulco hipocampal e fibras mielínicas aferentes |
 |
 |
 |
| Alveus | Fímbria | Plexo coróide |
 |
 |
 |
| Corpo geniculado lateral. Neurônios, axônios, lipofuscina. | Cauda do núcleo caudado. Neurônios, axônios | Área de infarto cortical. Córtex normal limítrofe |
 |
 |
 |
| Textos
: Estrutura do hipocampo, giro denteado, subículo,
conexões do hipocampo, esquema. |
 |
|
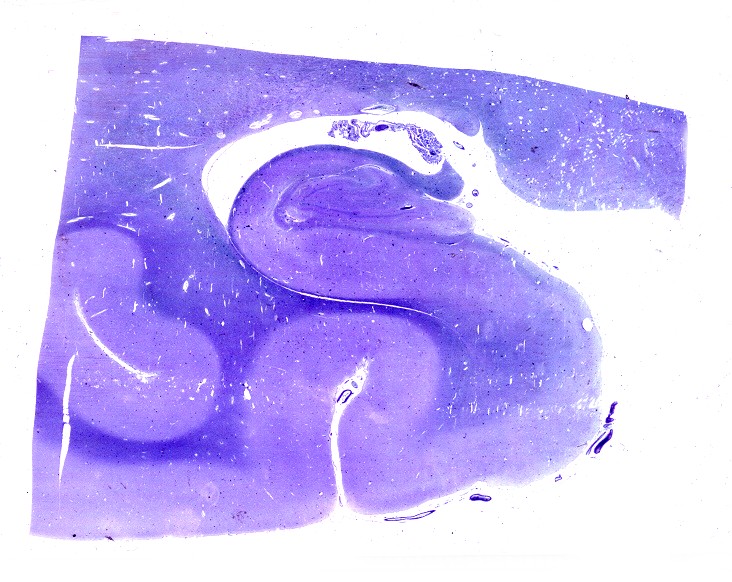
| LFB-Nissl.
Lâmina escaneada.
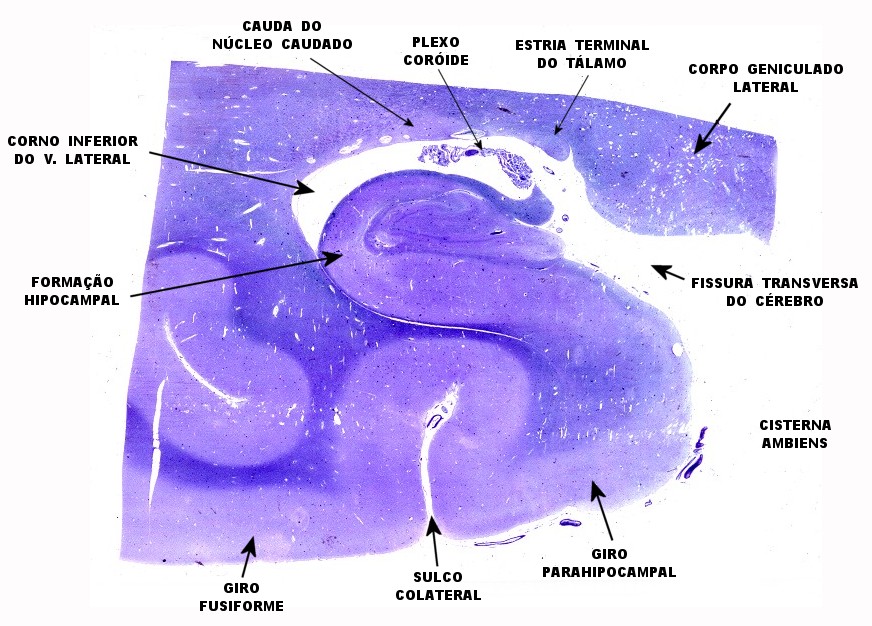
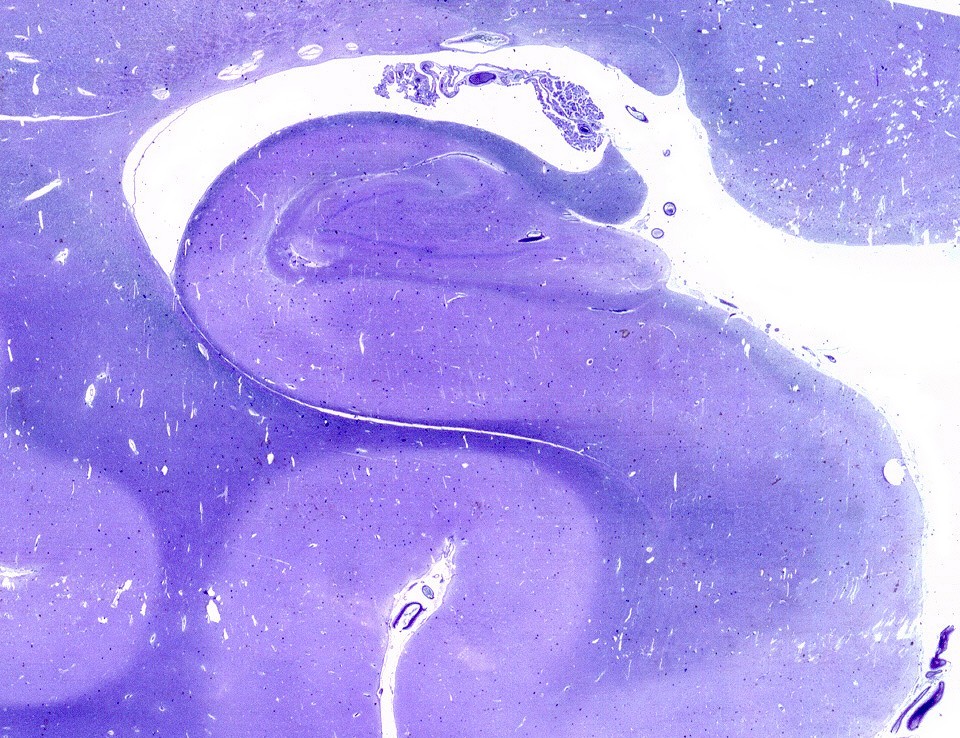
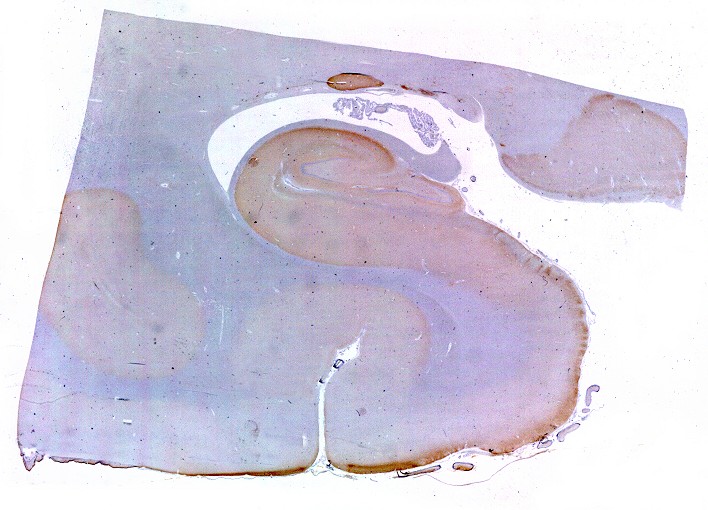
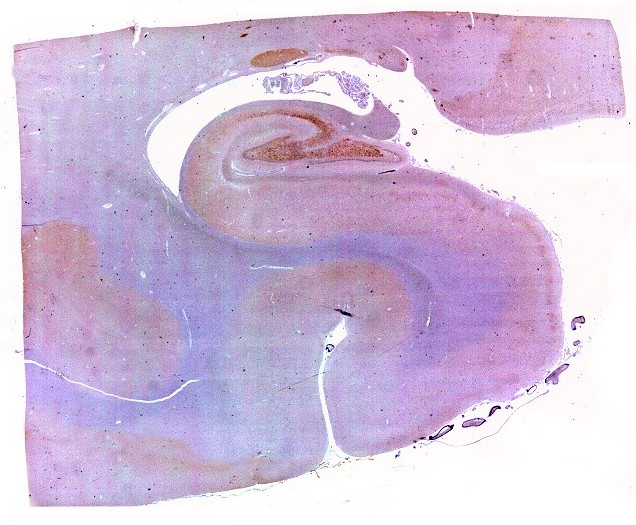
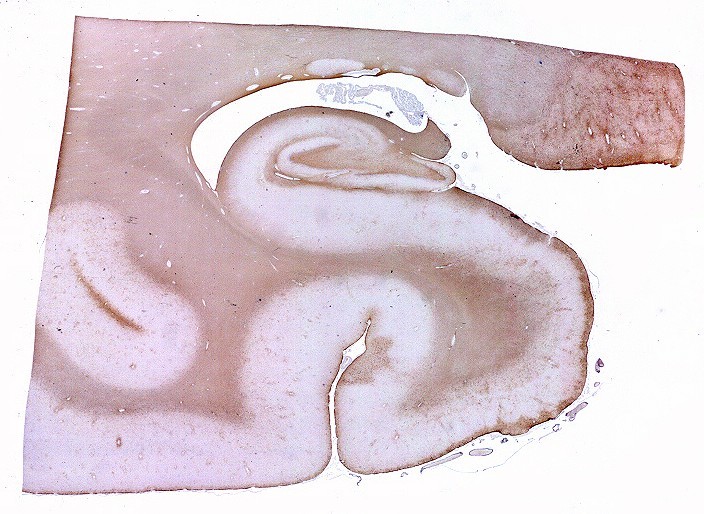
Mostra formação hipocampal, giro parahipocampal e seu vizinho, o giro fusiforme, e o corno inferior do ventrículo lateral. Substância branca em azul intenso, substância cinzenta em tons mais claros. Para procedimento técnico do LFB-Nissl, clique. Lâmina escaneada em 600 dpi (dots per inch). Foto seguinte com 1200 dpi mostra setores do hipocampo. Para texto sobre anatomia e texto e esquema sobre conexões do hipocampo, clique. |
 |
 |
 |
 |
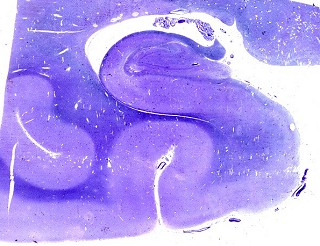
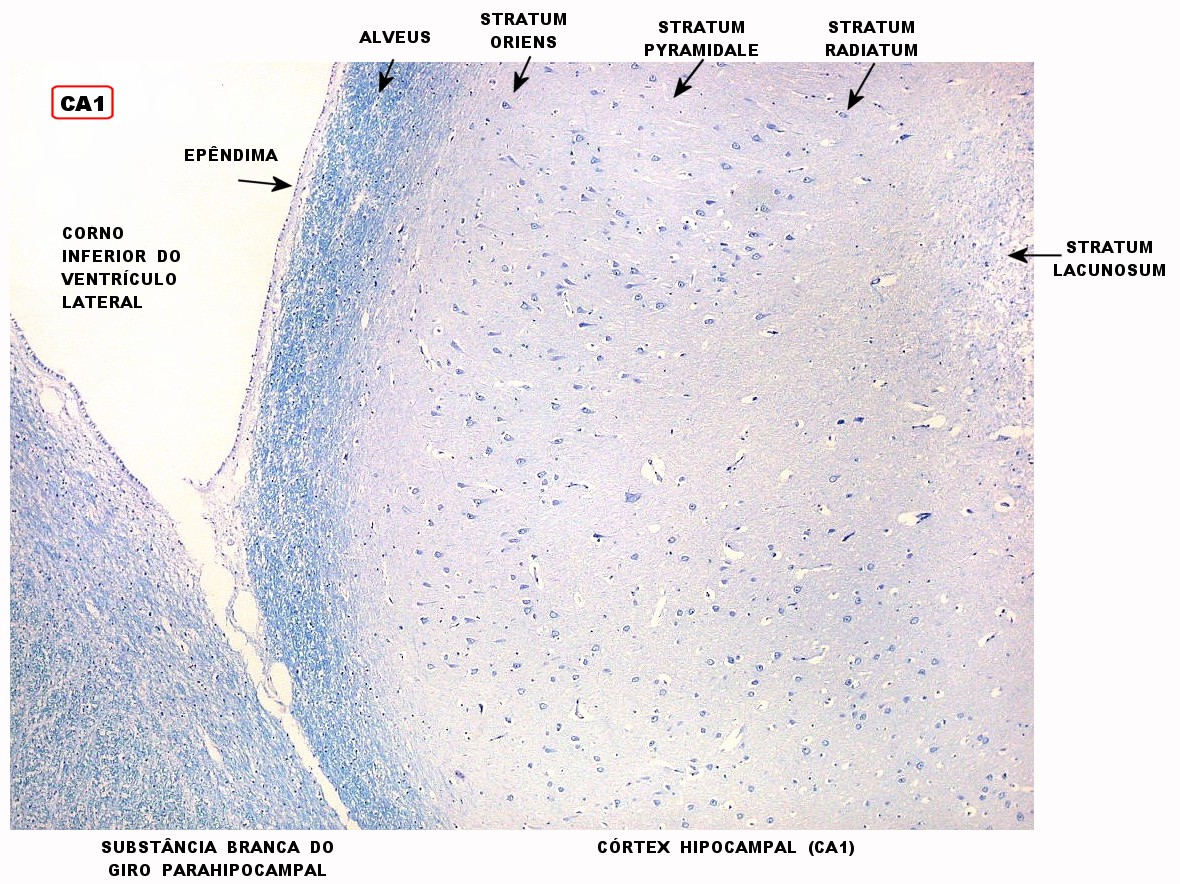
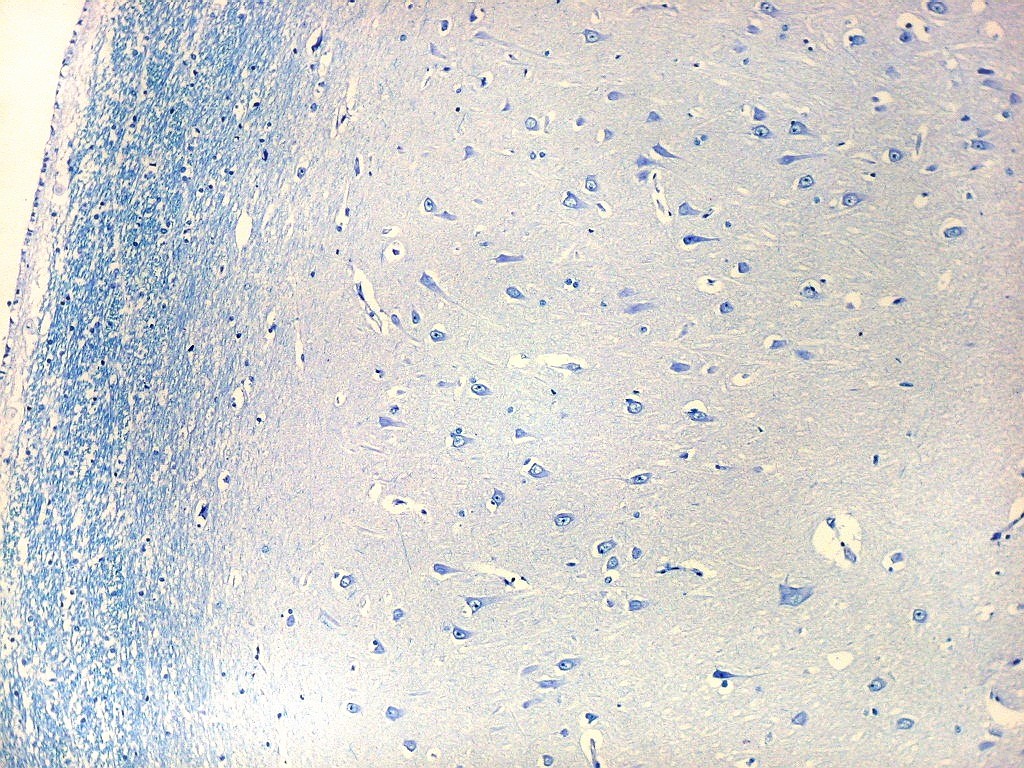
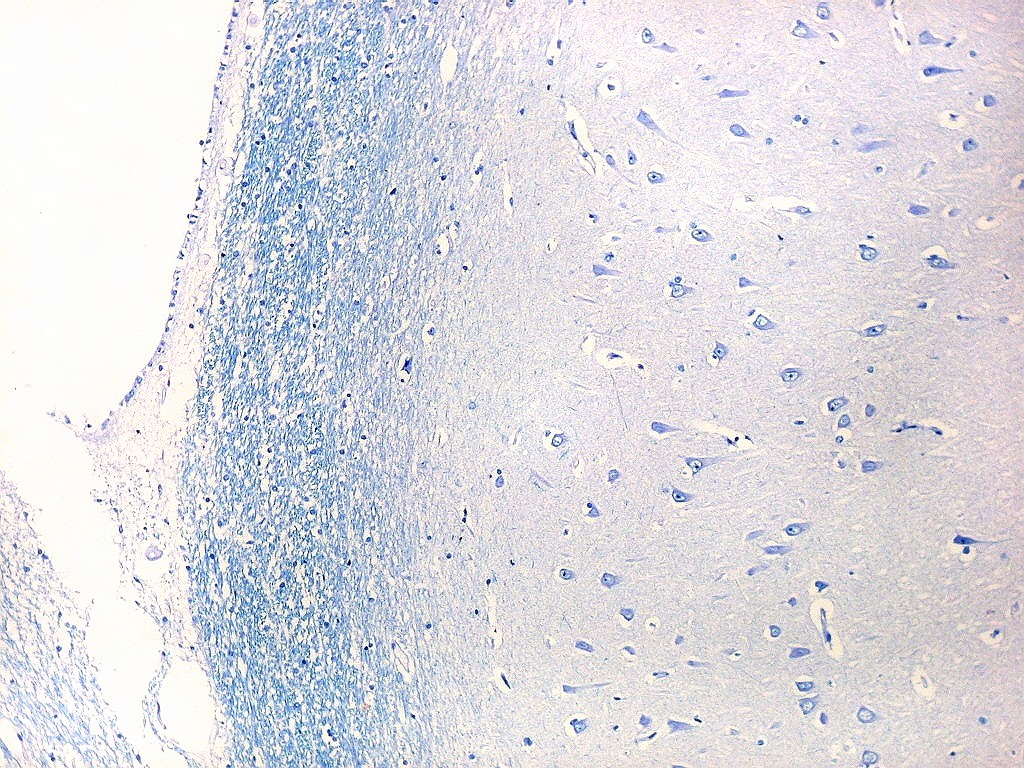
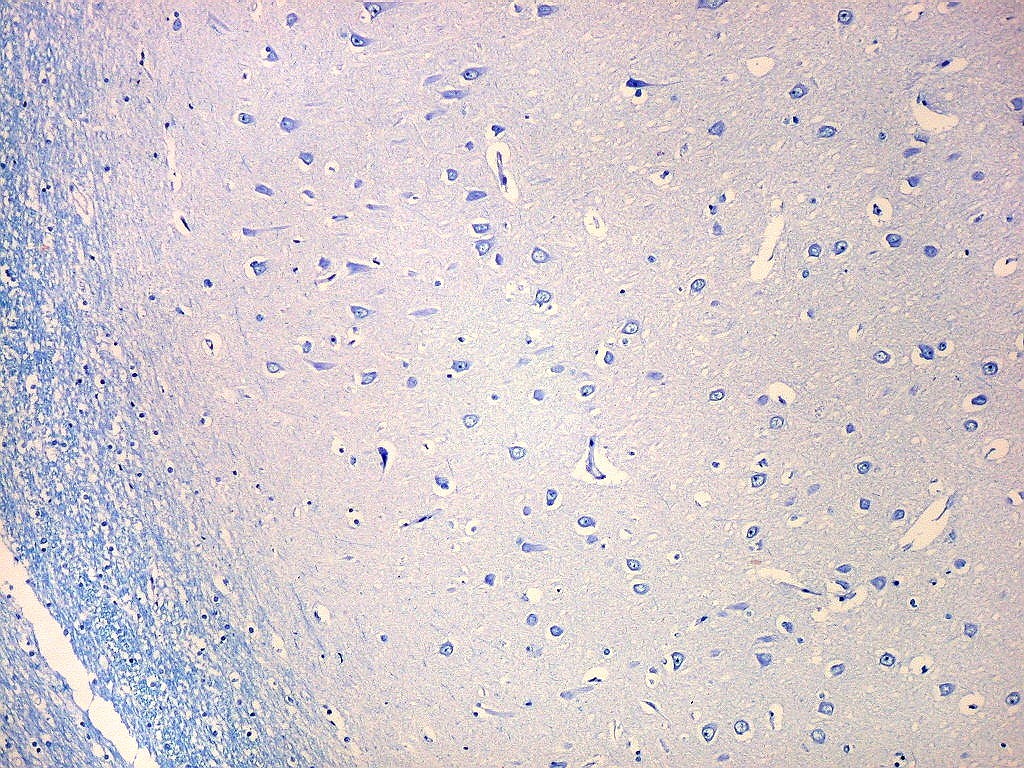
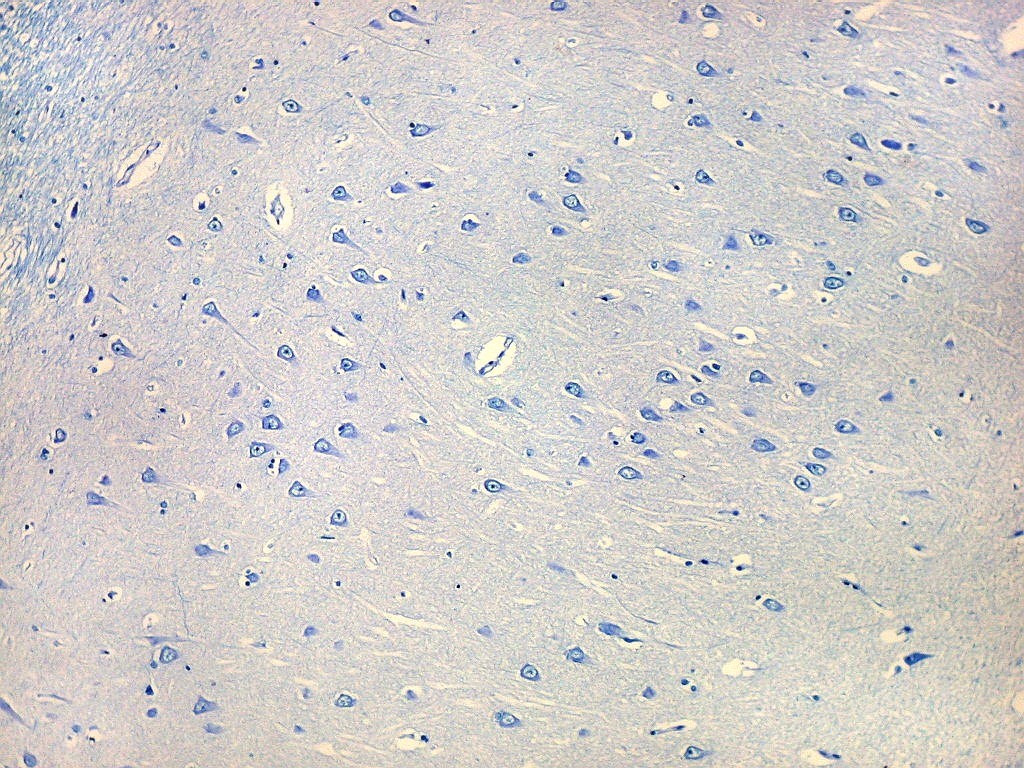
| Setor CA1. Aqui mostrado em sua porção média, próximo ao ângulo do corno inferior do ventrículo lateral, mostra neurônios pequenos, triangulares, dispersos em abundante neurópilo. Dendritos apicais são dirigidos internamente (à direita na foto), axônios em sentido oposto, em direção ao alveus (à esquerda). Como o alveus é uma camada de substância branca, aparece em azul mais intenso devido à abundância de axônios mielínicos. |
 |
 |
 |
 |
 |
 |
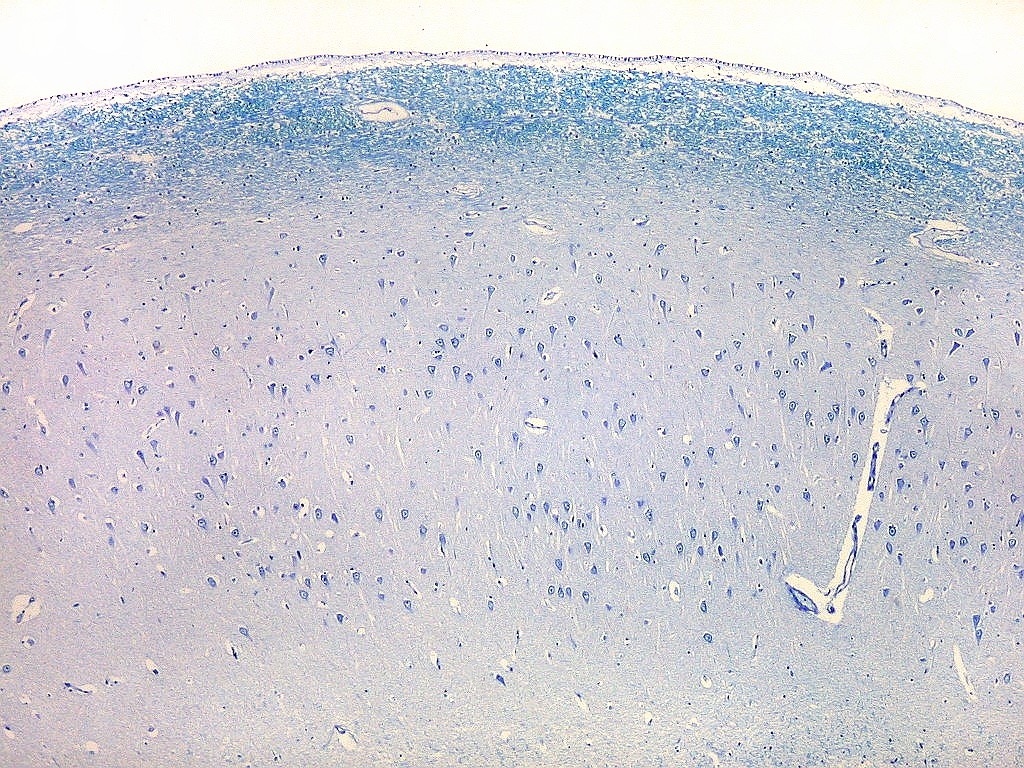
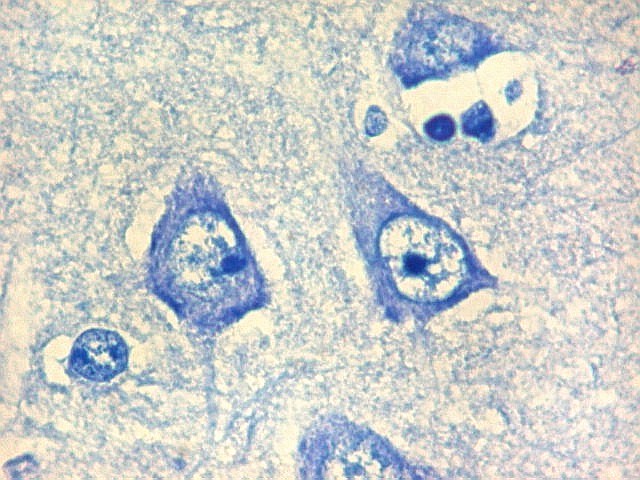
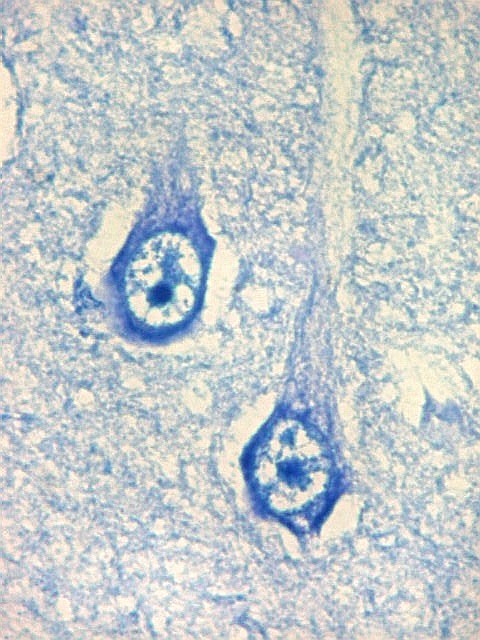
| Setor CA1. Aqui na porção mais alta, próxima à transição para setor CA2. Em cima, epêndima do ventrículo e alveus (camada de fibras mielínicas em azul turquesa). Neurônios piramidais com dendritos apicais para baixo (na foto). Nos detalhes feitos com objetiva de imersão, abaixo, os neurônios foram rodados 180 graus, ficando com os dendritos apicais para cima. |
 |
 |
|
 |
 |
 |
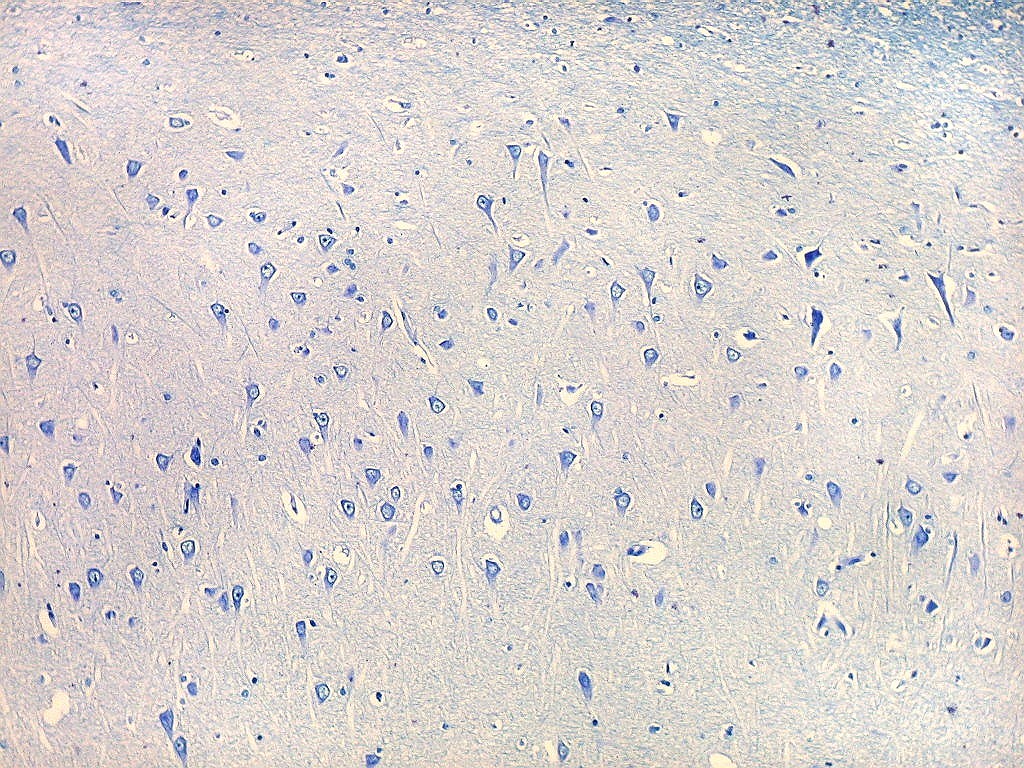
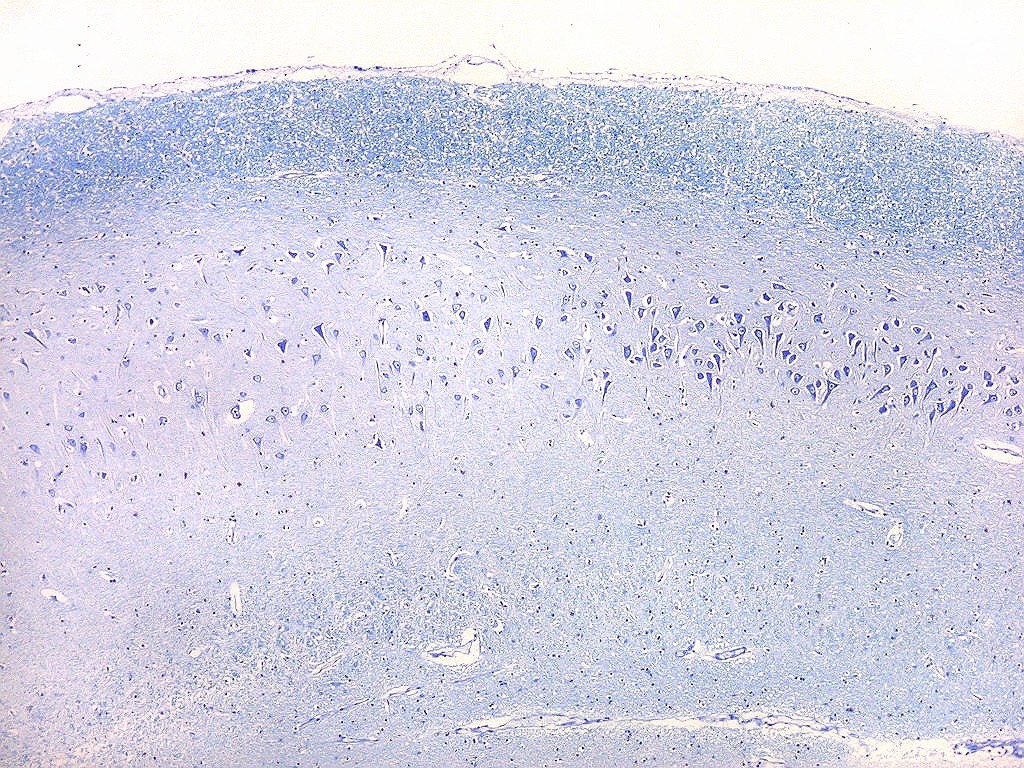
| Transição do setor CA1 para CA2. A transição é relativamente abrupta, com os neurônios piramidais em CA1 (à esquerda) mais dispersos e espaçados, em contraste com os de CA2, mais condensados e ocupando uma faixa mais estreita. |
 |
 |
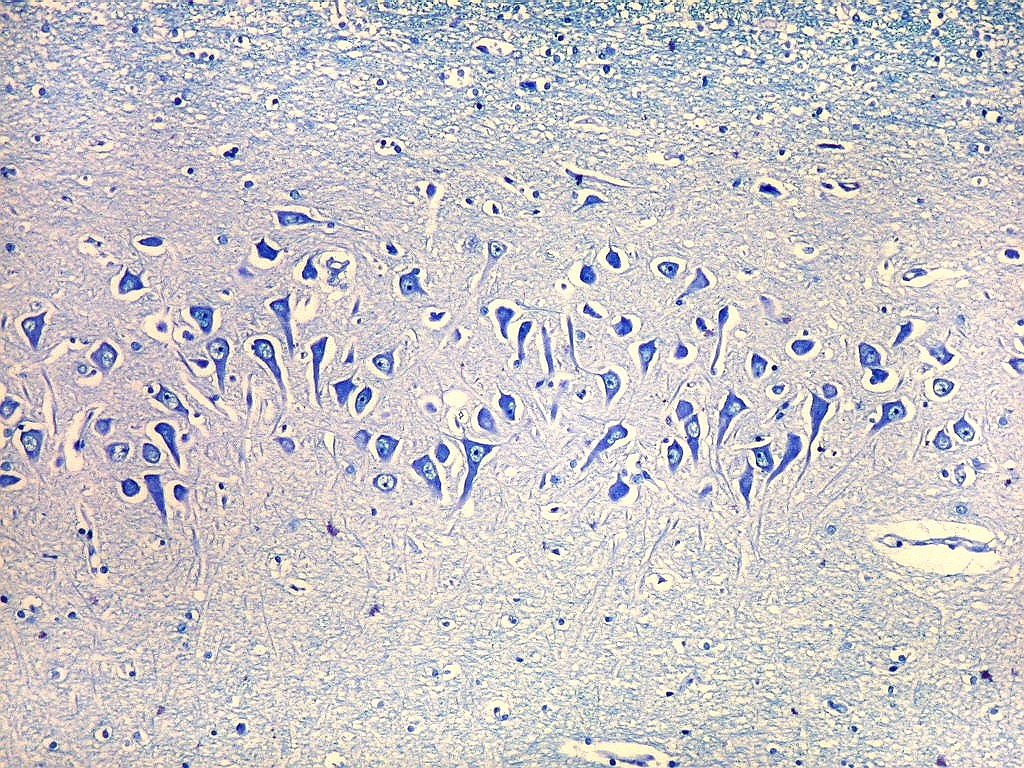
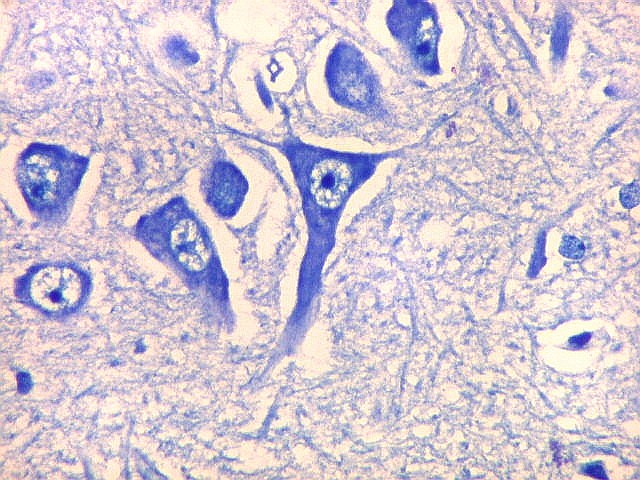
| Setor CA2. Menos extenso que CA1, neurônios piramidais maiores e mais densamente agrupados, em disposição radial paralela semelhante à de CA1. |
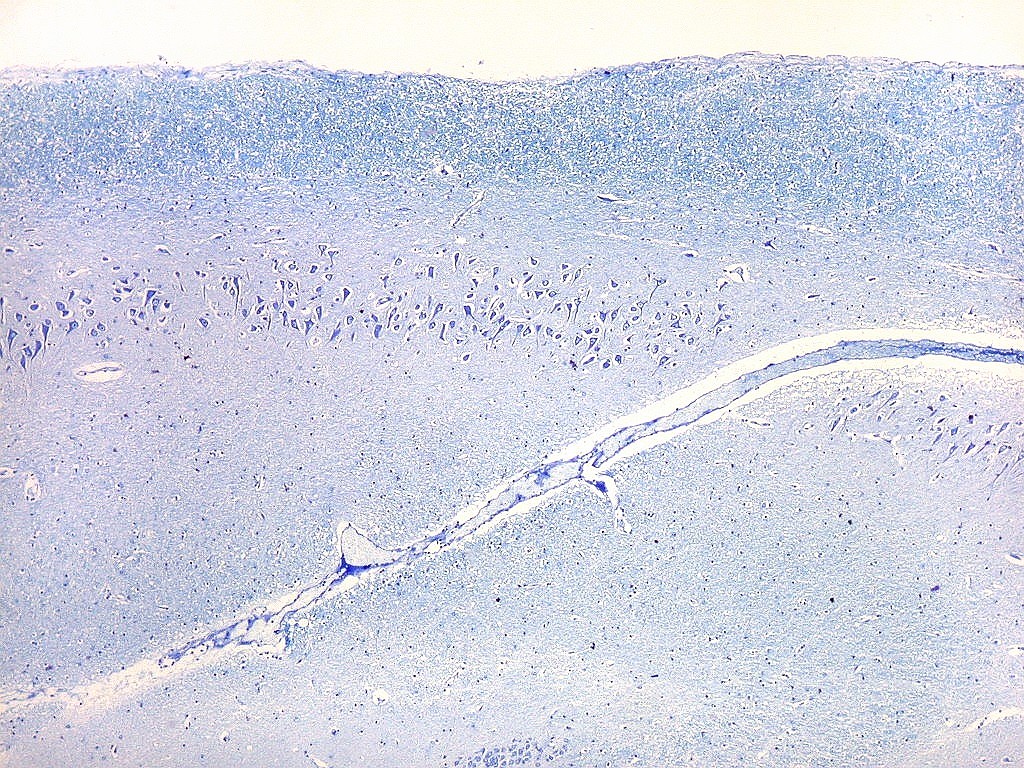 |
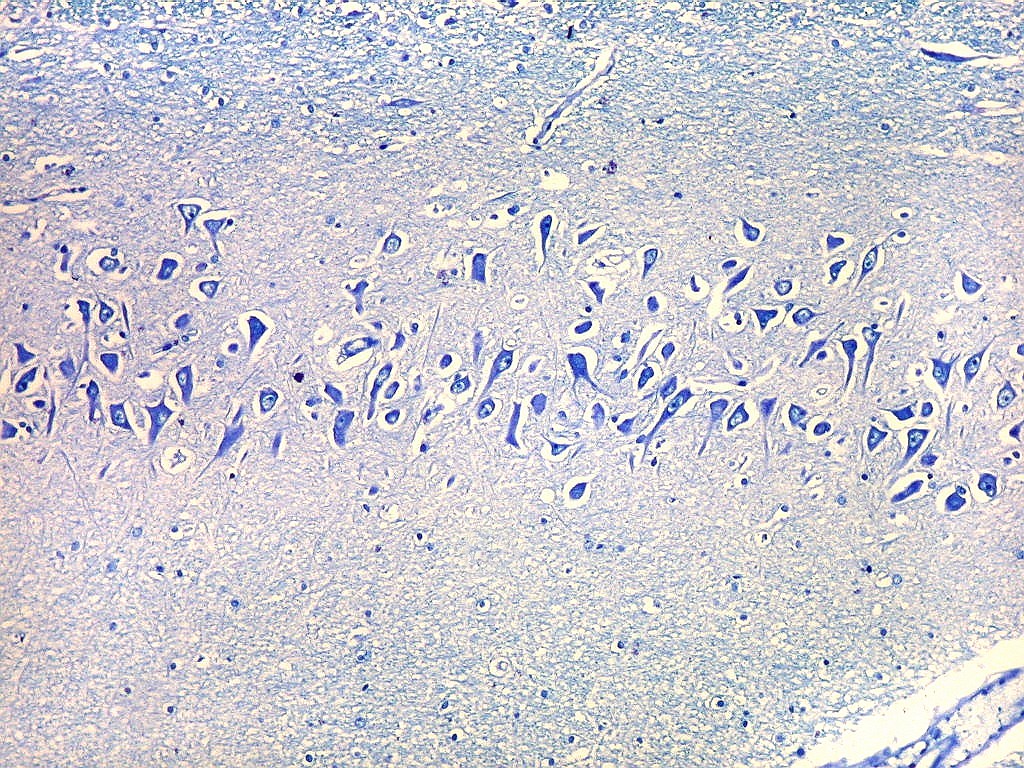 |
 |
|
 |
 |
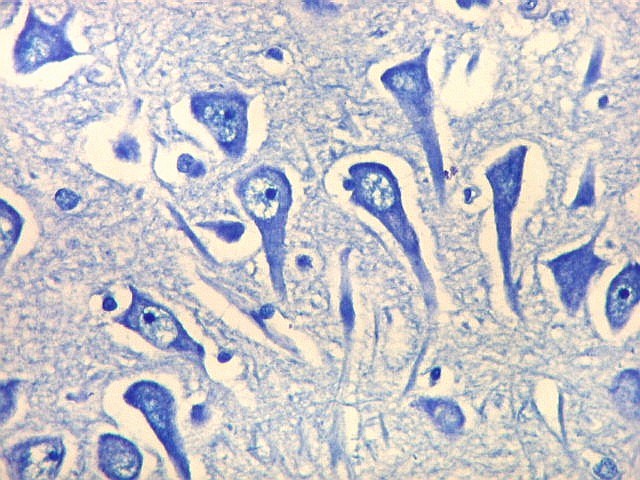 |
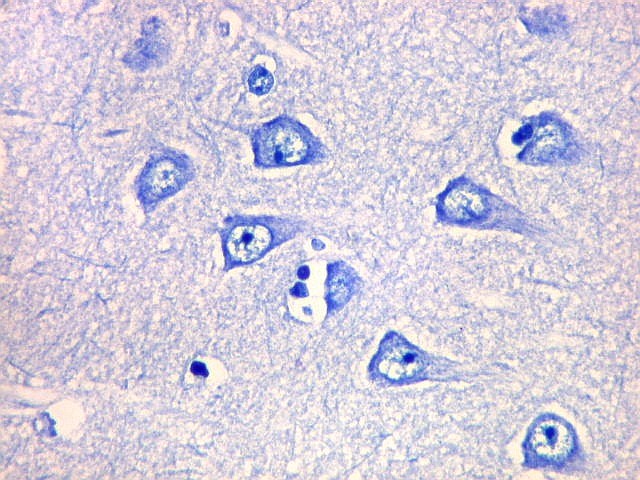
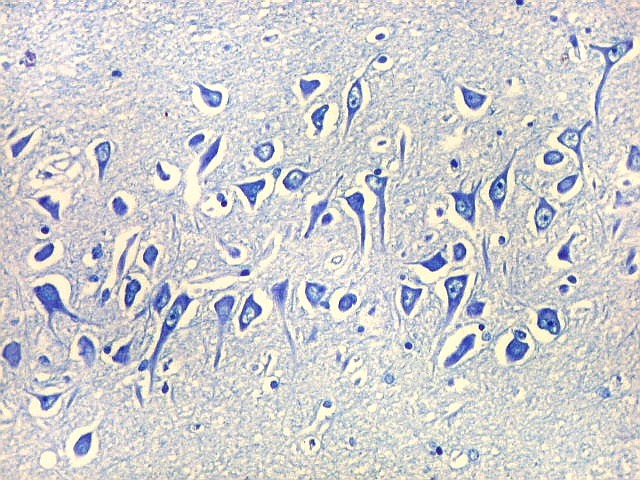
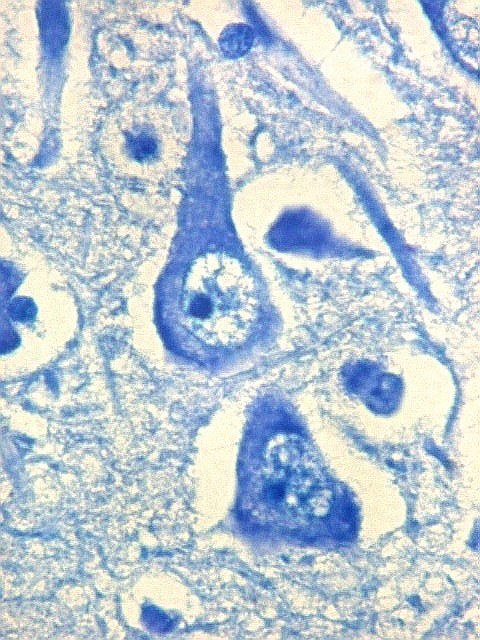
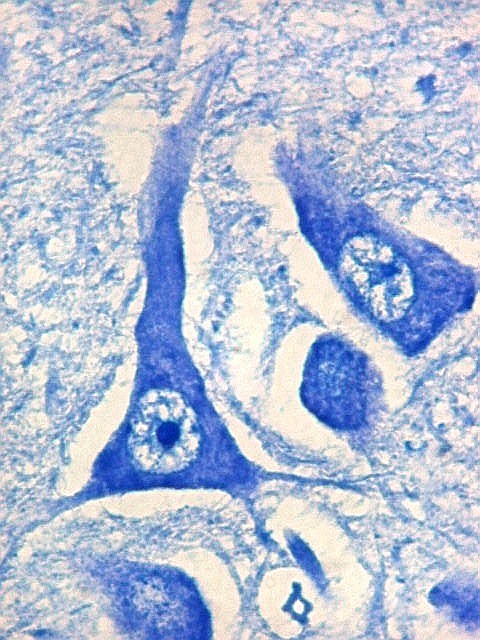
| CA2. Nas fotos abaixo, neurônios piramidais fotografados com objetiva de imersão e apresentados com seus dendritos apicais para cima (rodados 180 graus em relação à posição anatômica). | |
 |
 |
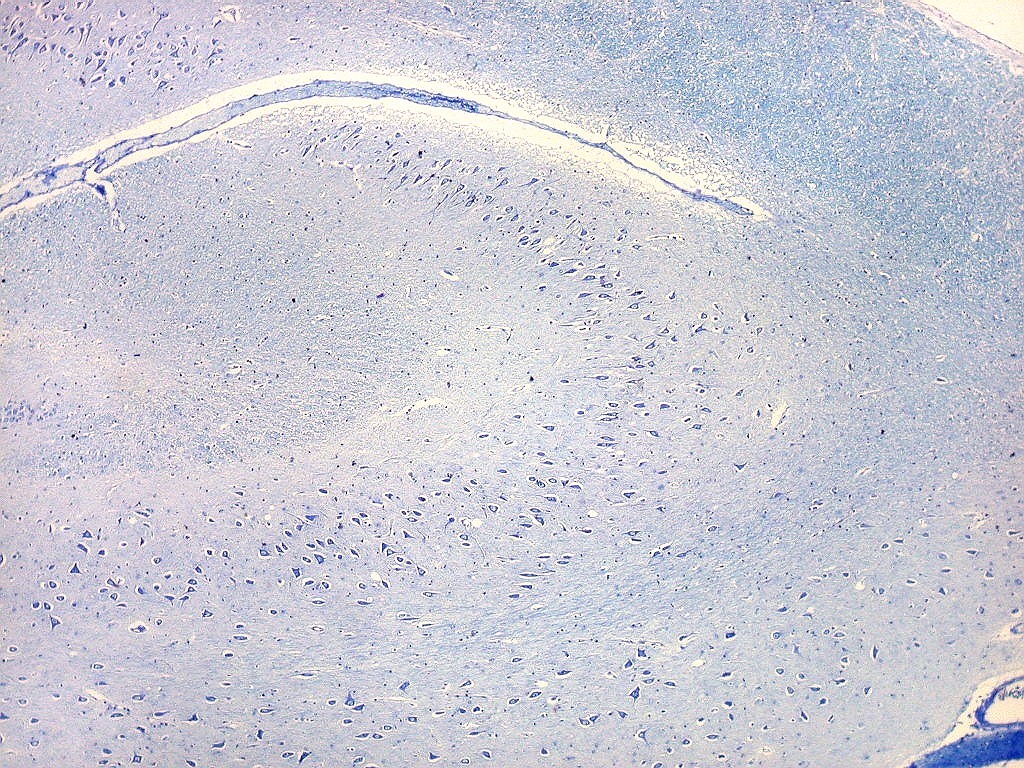
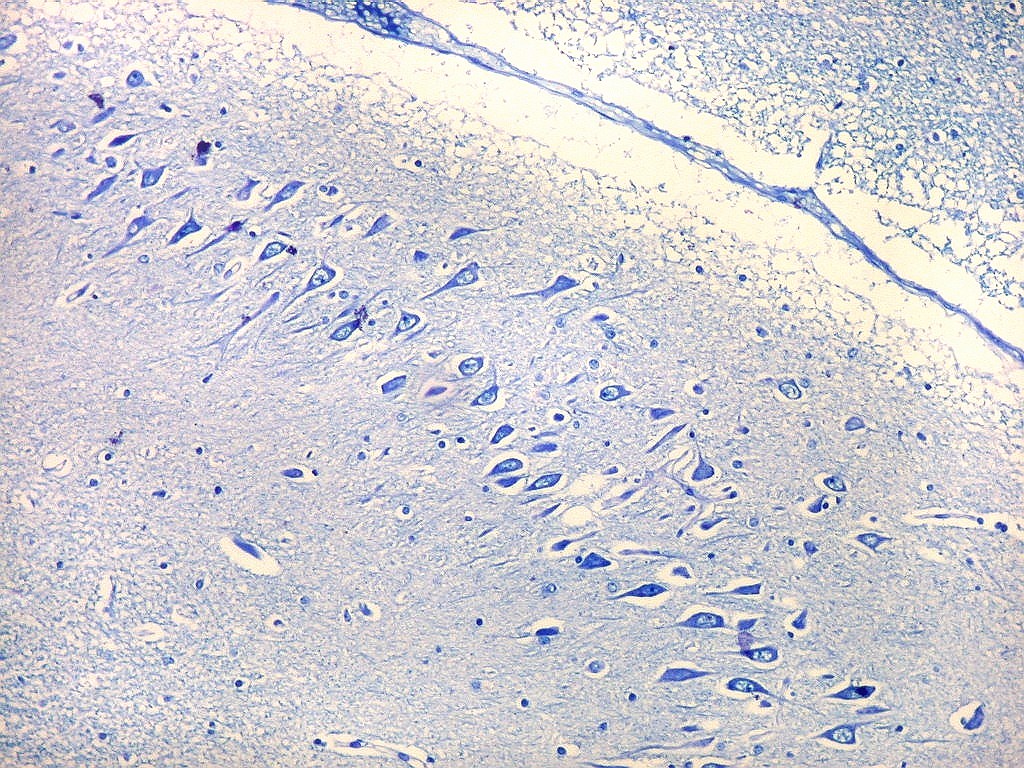
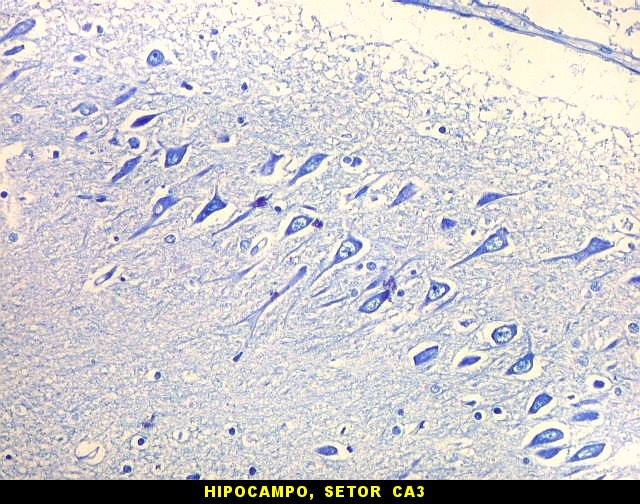
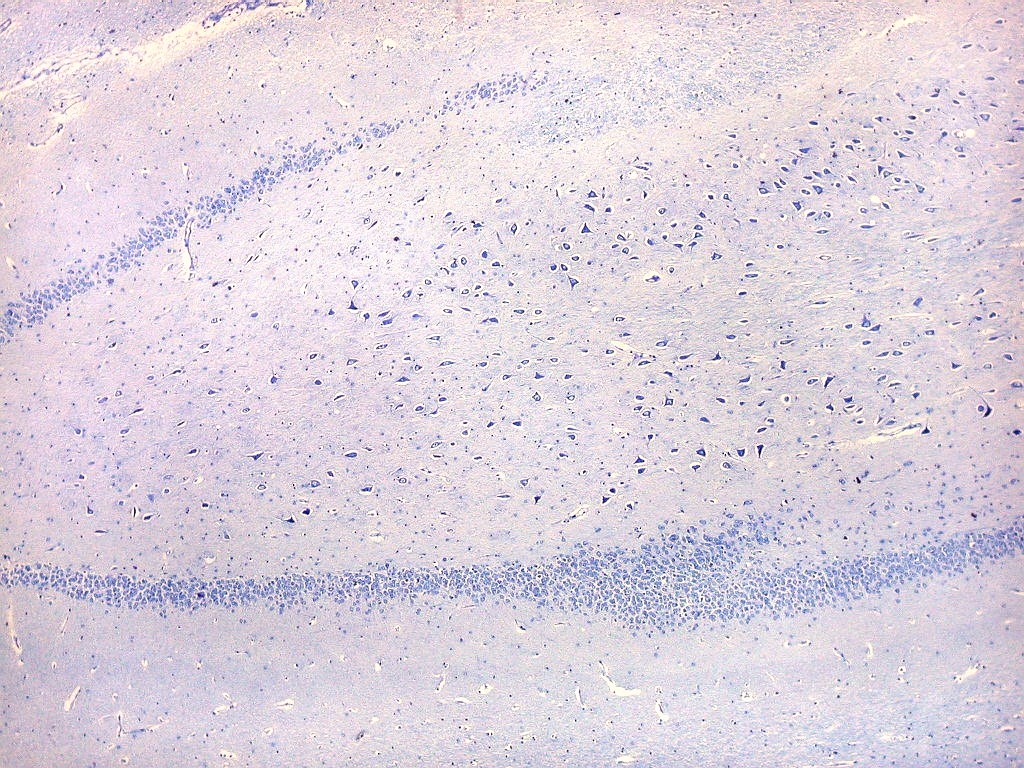
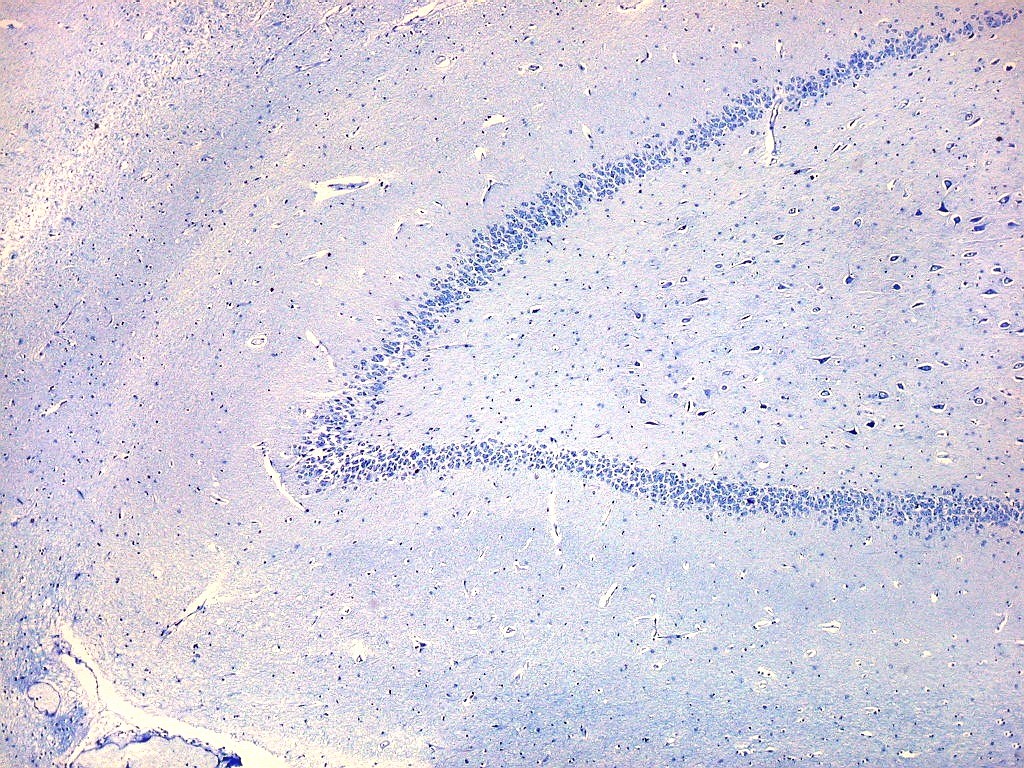
| Setor CA3. Continua o setor CA2 com a curvatura da camada piramidal em direção a CA4 (para baixo na foto). CA4 é o setor do hipocampo envolvido pelo giro denteado. A disposição radiada dos neurônios em CA3 é como a de CA2, mas vão se tornando mais dispersos e menos orientados à medida que fazem transição para CA4. |
 |
 |
 |
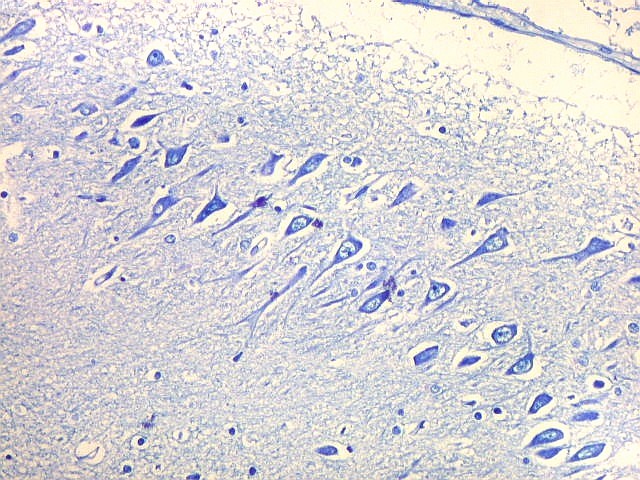
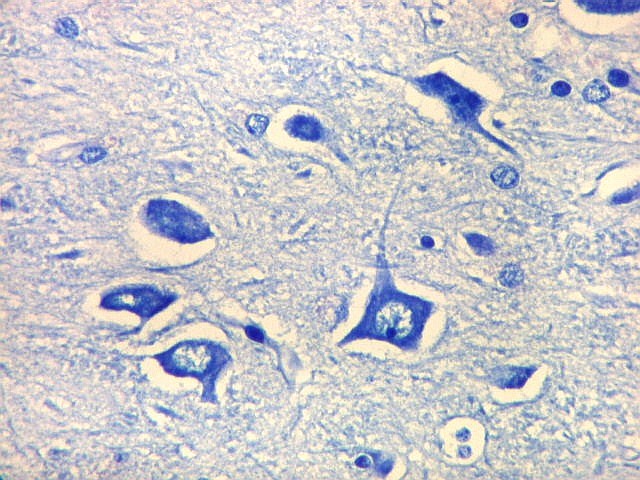
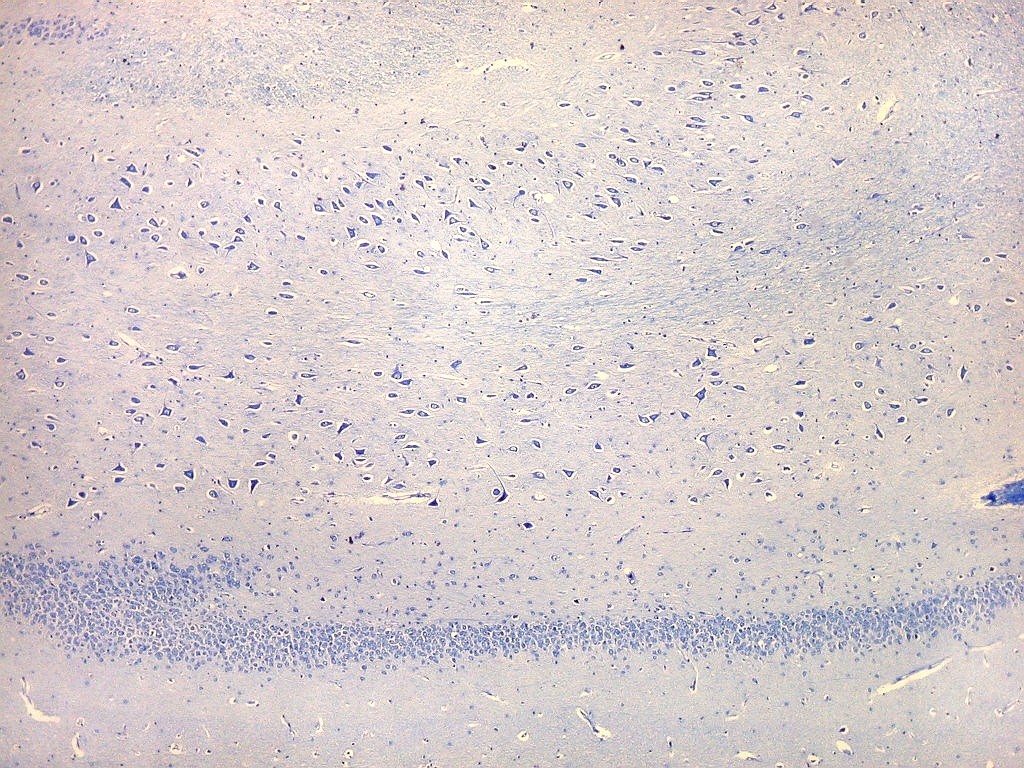
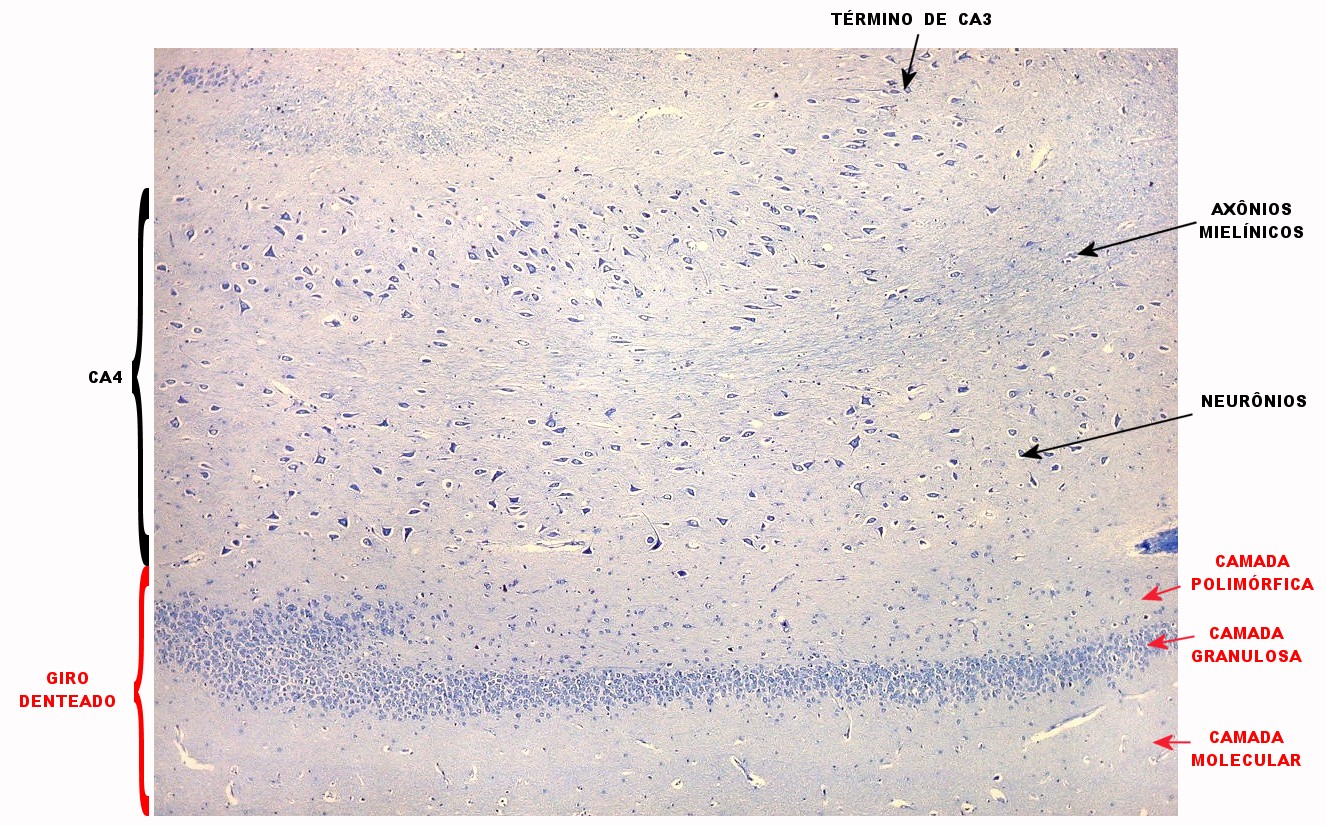
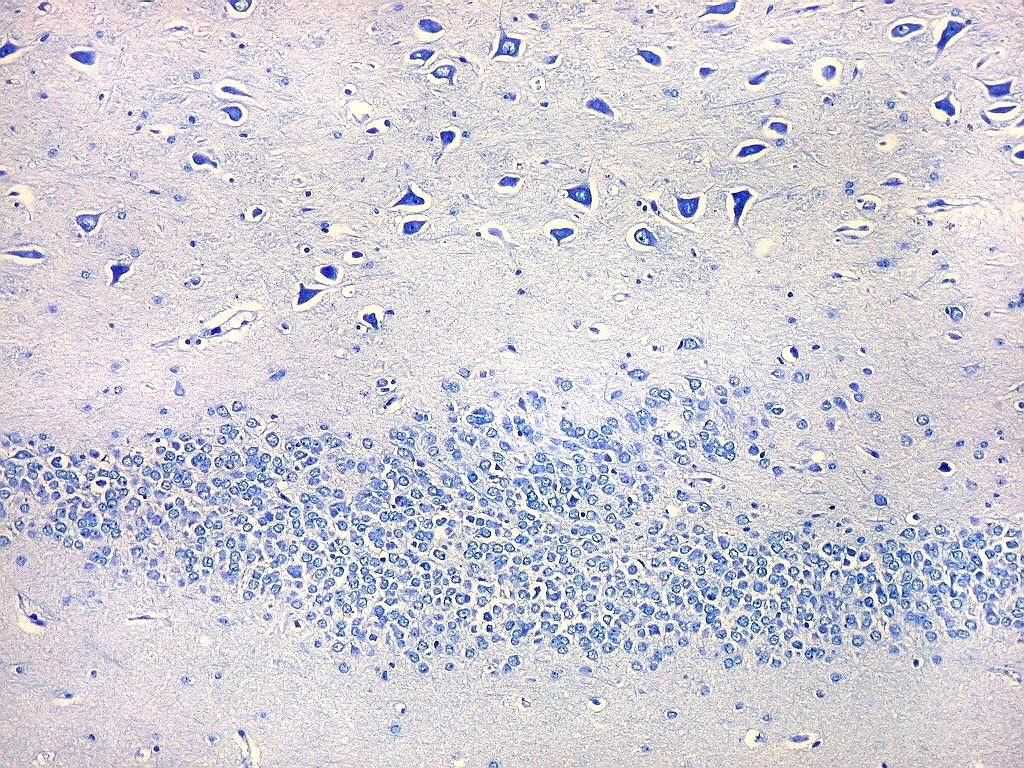
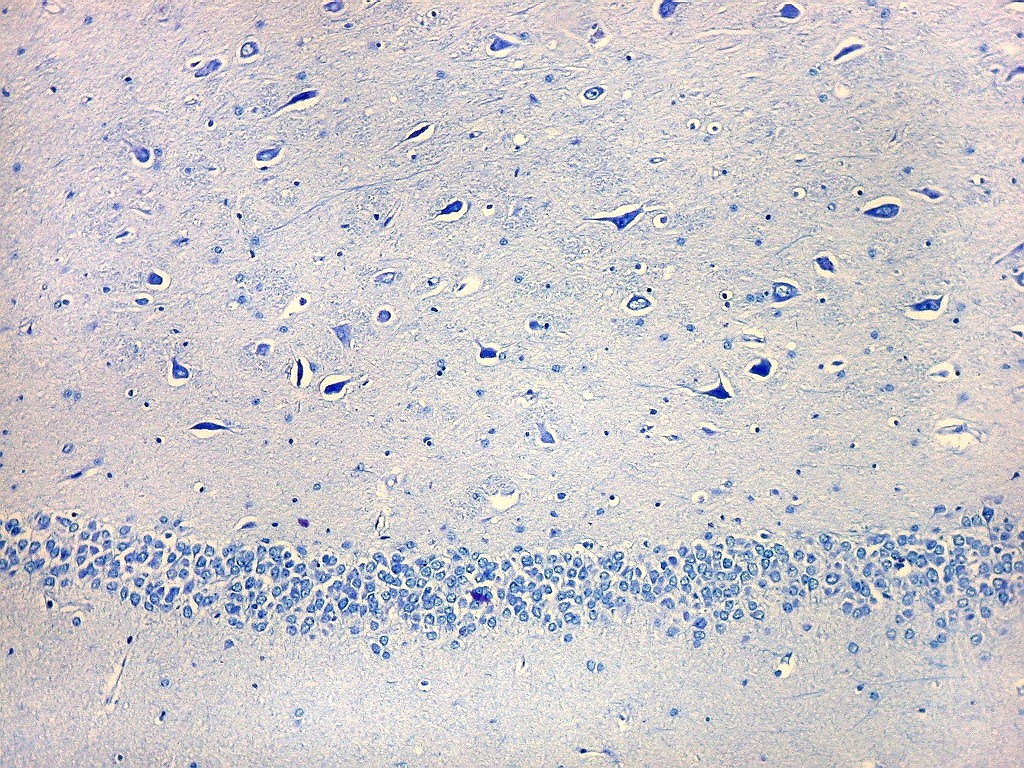
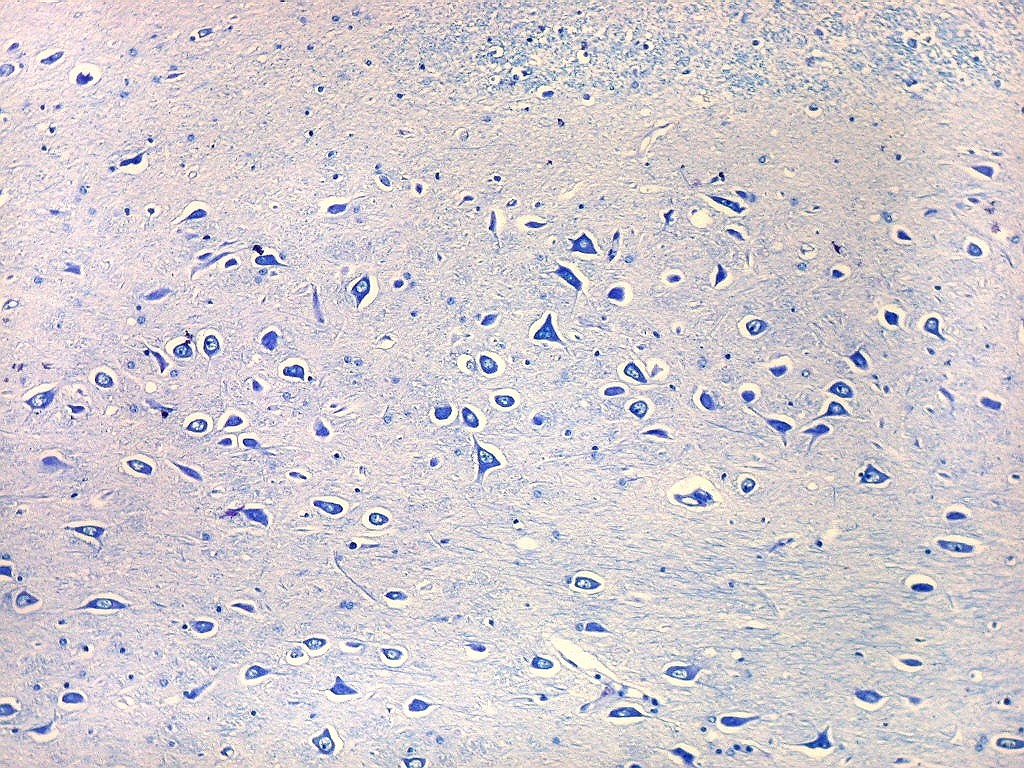
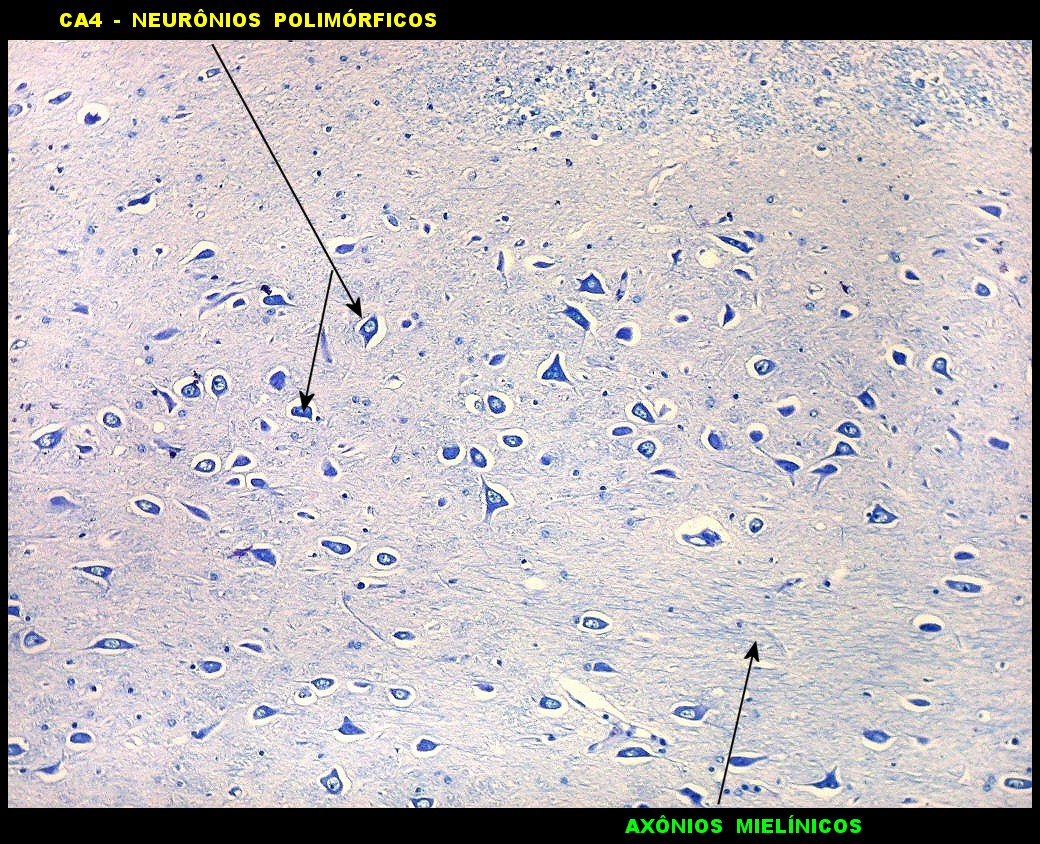
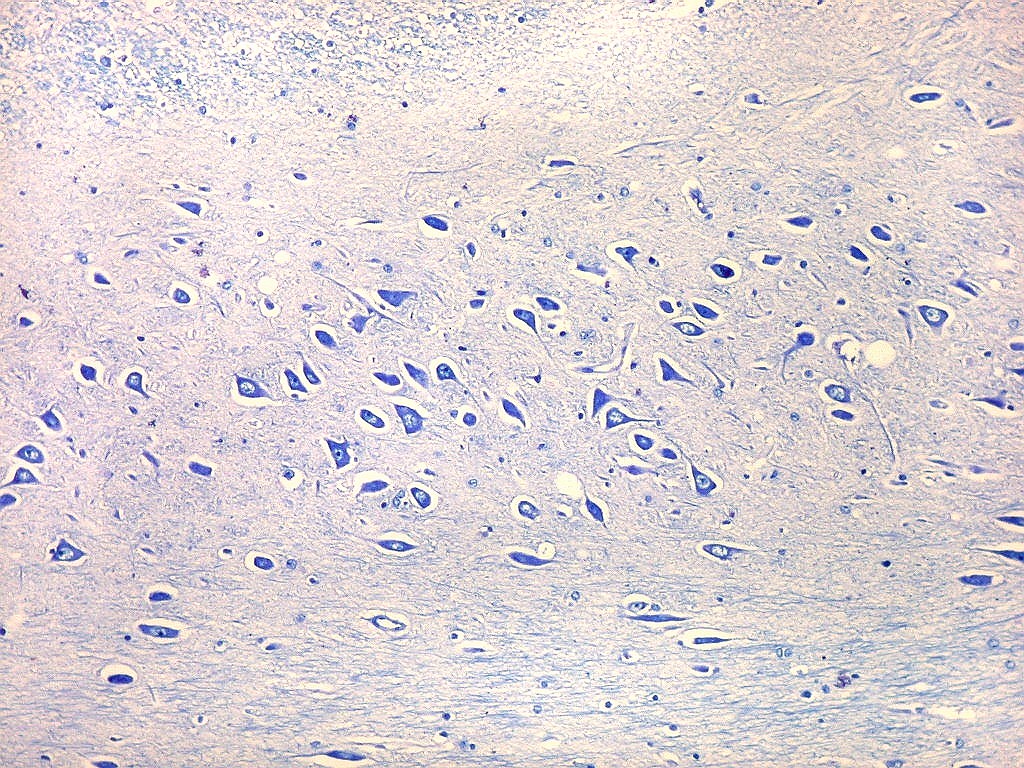
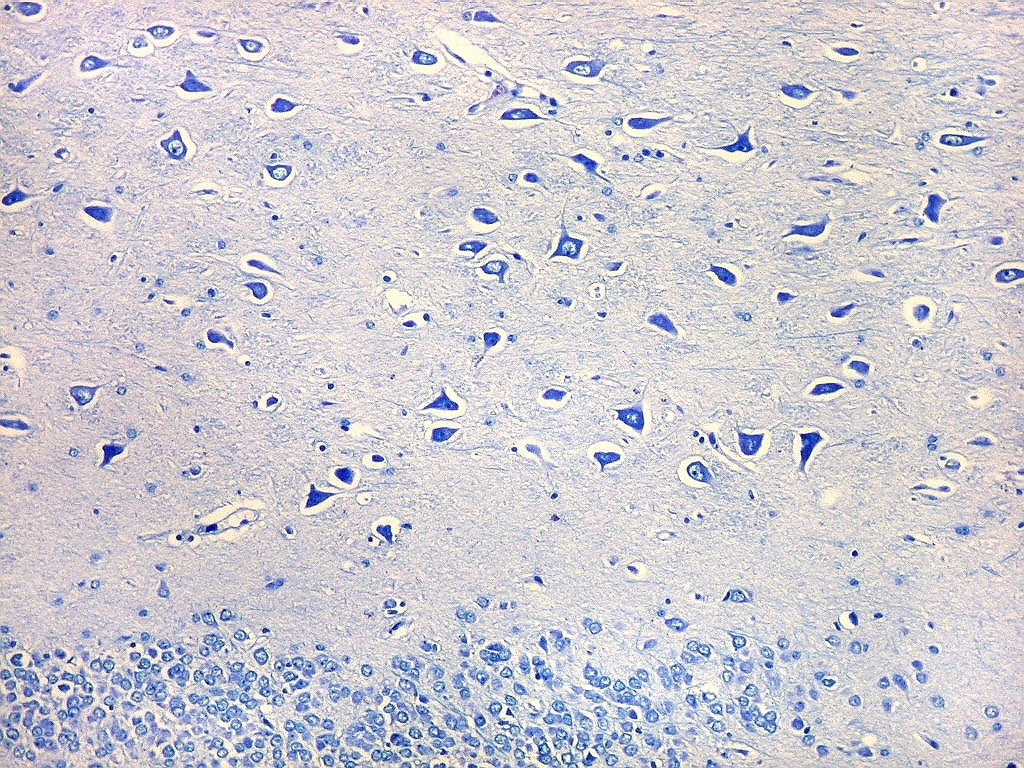
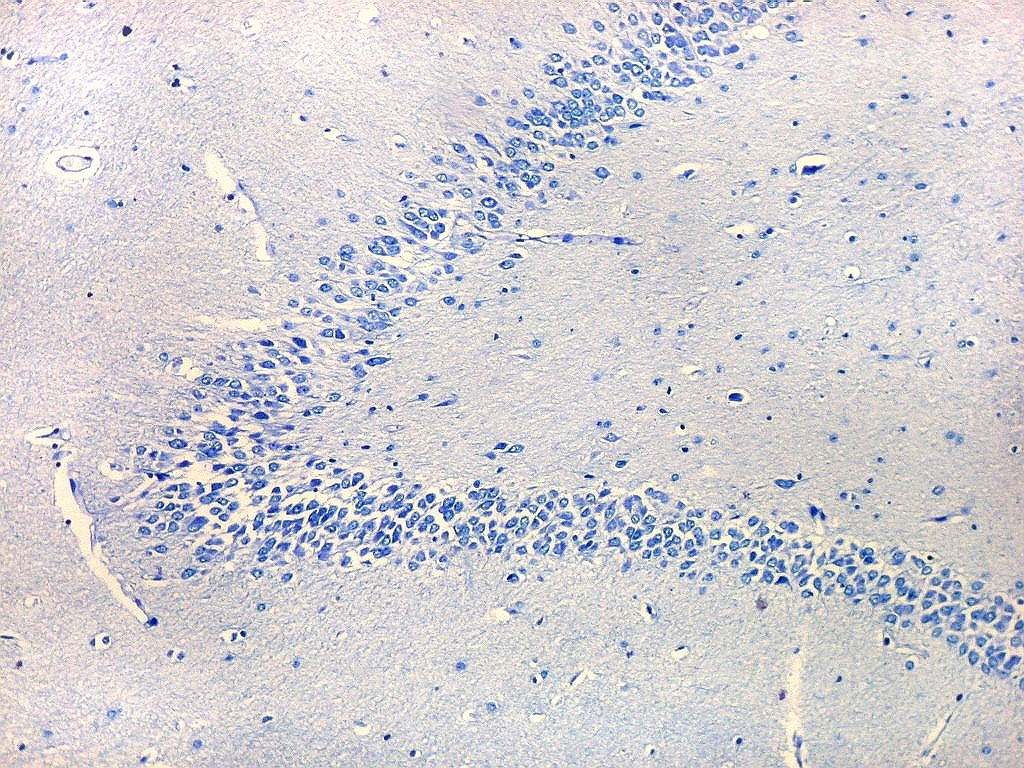
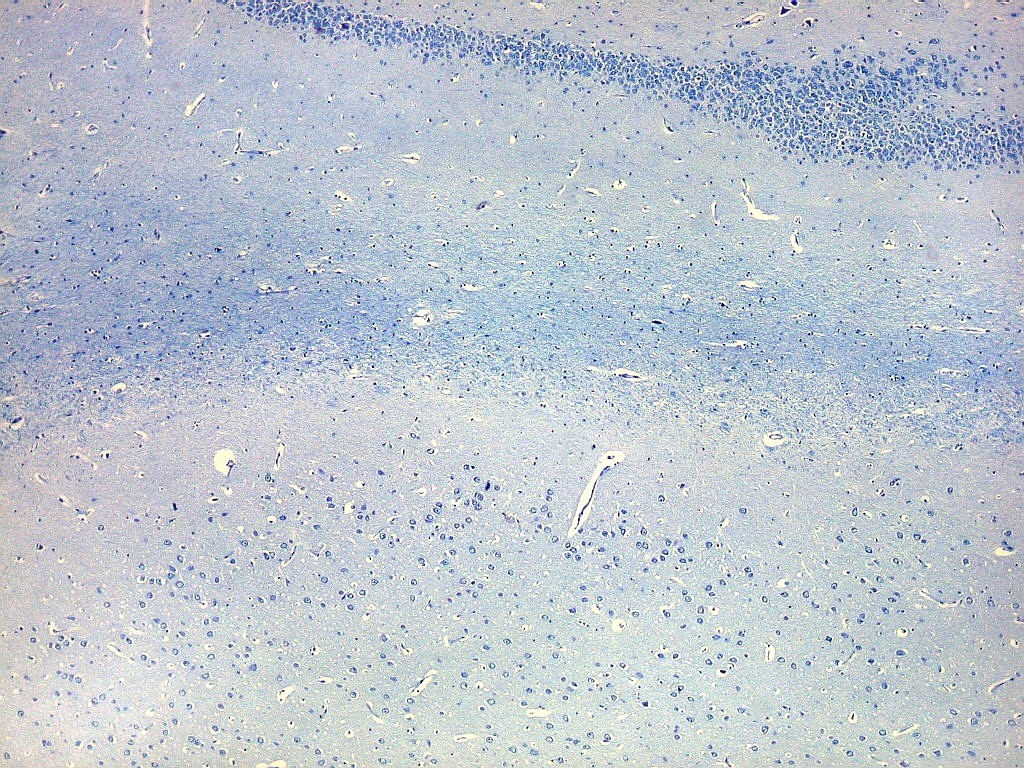
| Setor CA4 e giro denteado. Em CA4, neurônios perdem a orientação radial paralela vista nos 3 setores acima. As células são polimórficas, algumas com contorno triangular, outras mais ovaladas e outras multipolares. Os pequenos neurônios granulosos do giro denteado circundam o setor como a cerca de um curral. |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
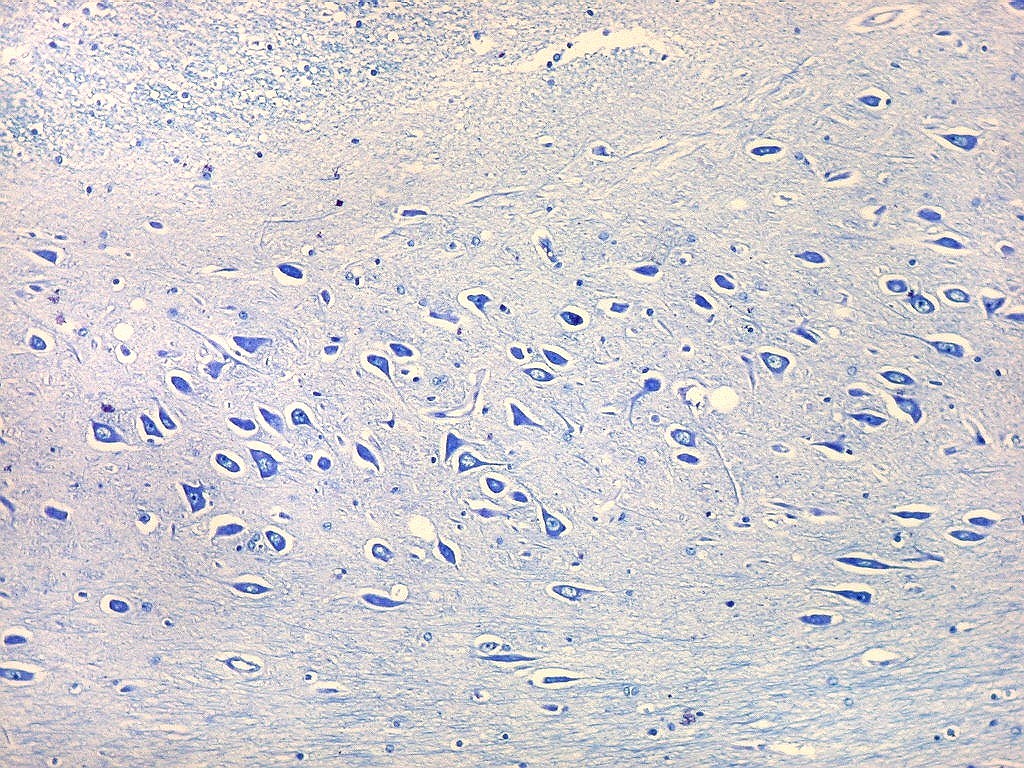 |
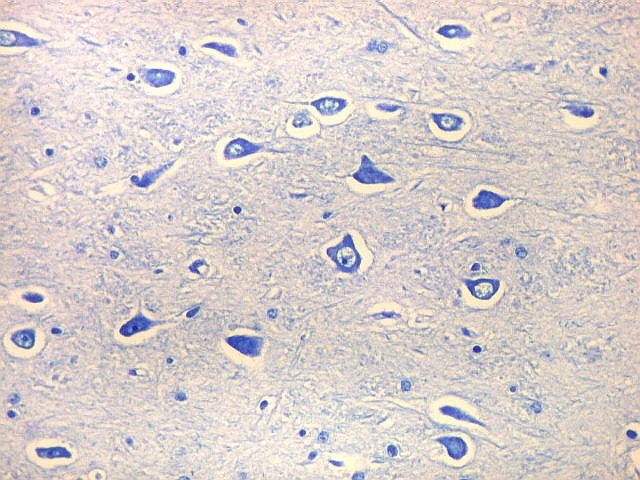
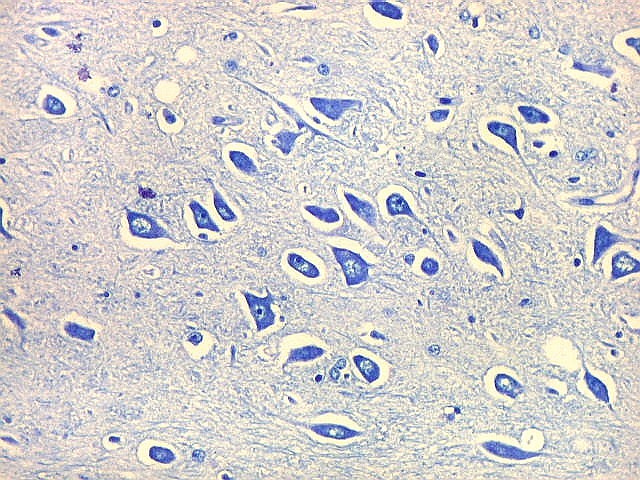
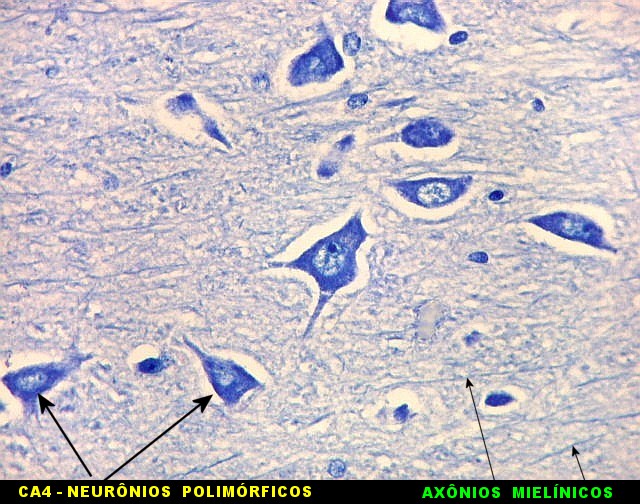
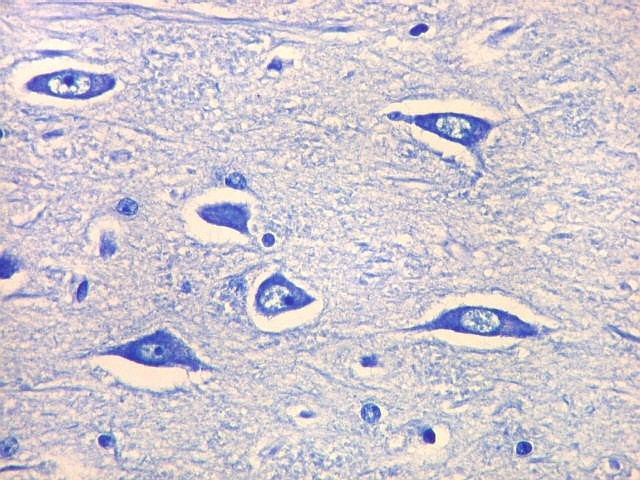
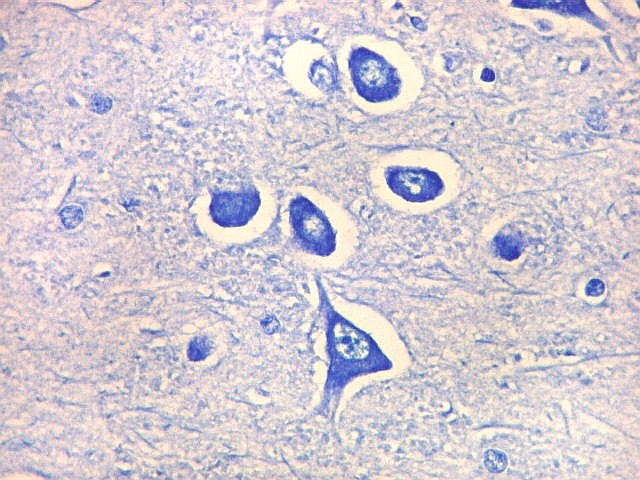
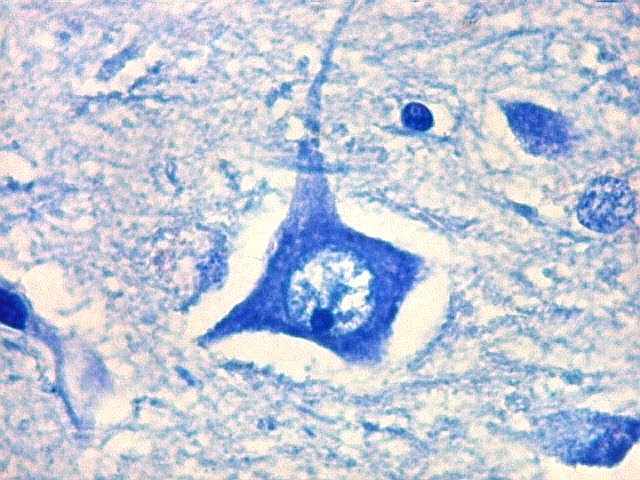
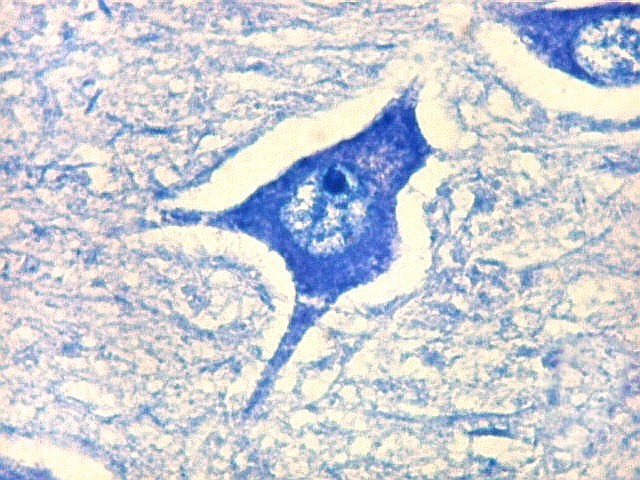
| CA4. Detalhes dos neurônios polimórficos do setor CA4. Os axônios dos neurônios granulosos que se distribuem entre eles não são visíveis nesta preparação por LFB-Nissl, mas podem ser observados na imunohistoquímica para cromogranina. Axônios mielínicos espessos podem ser vistos cursando entre os neurônios maiores. | |
 |
 |
 |
|
 |
 |
 |
 |
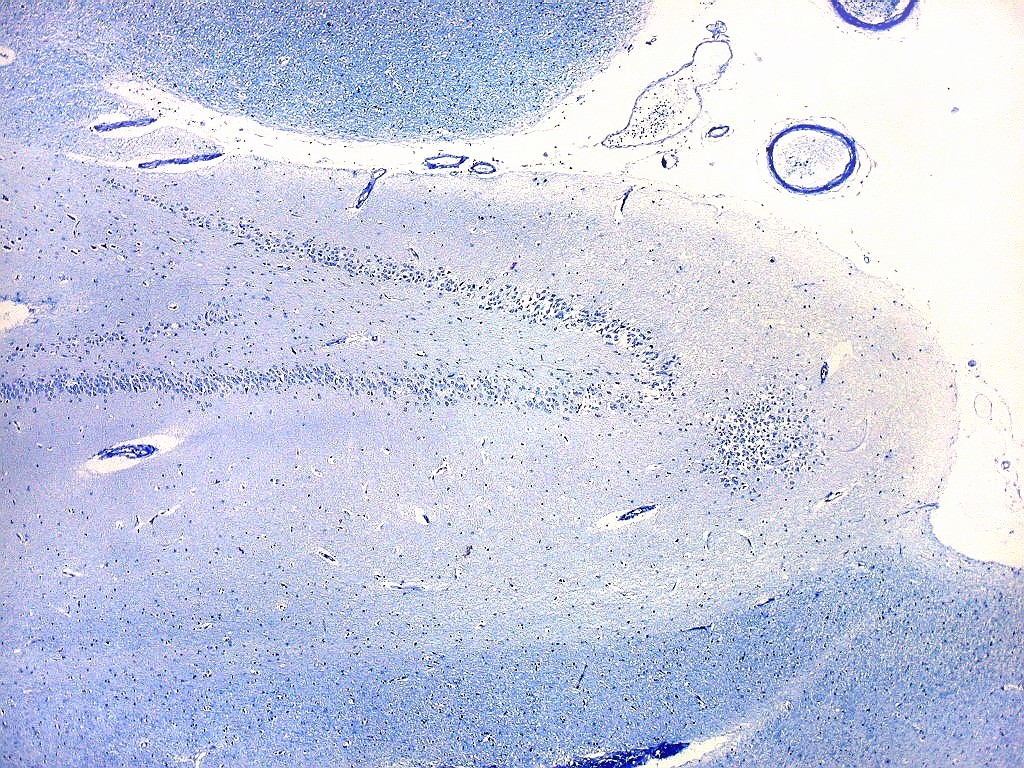
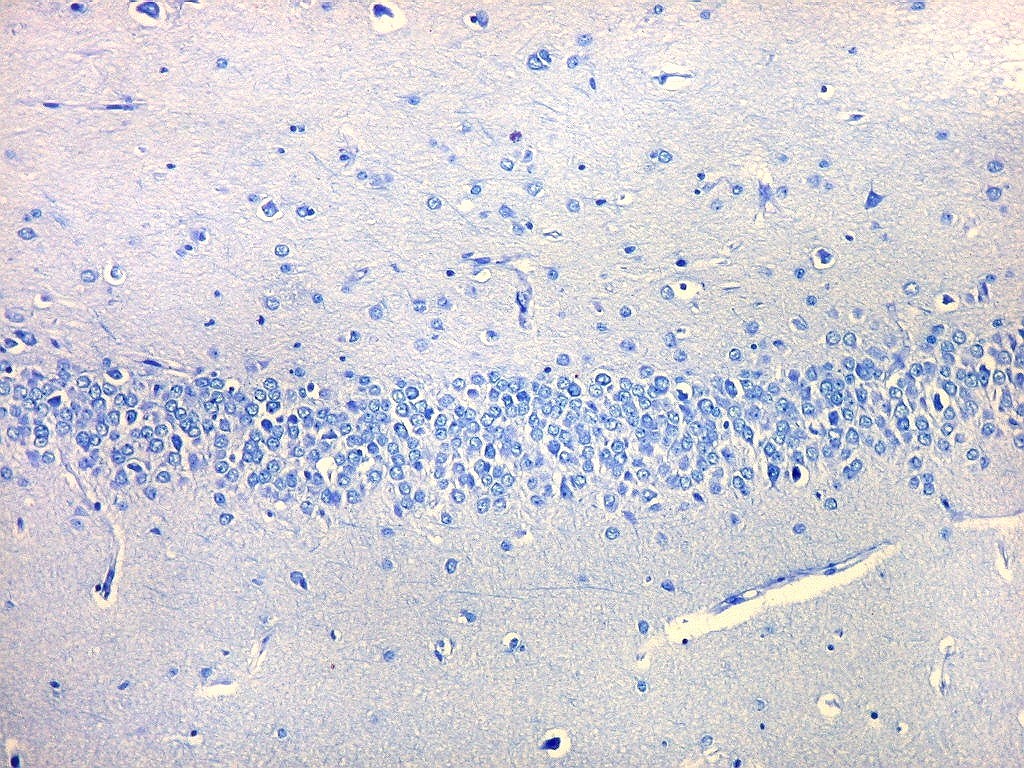
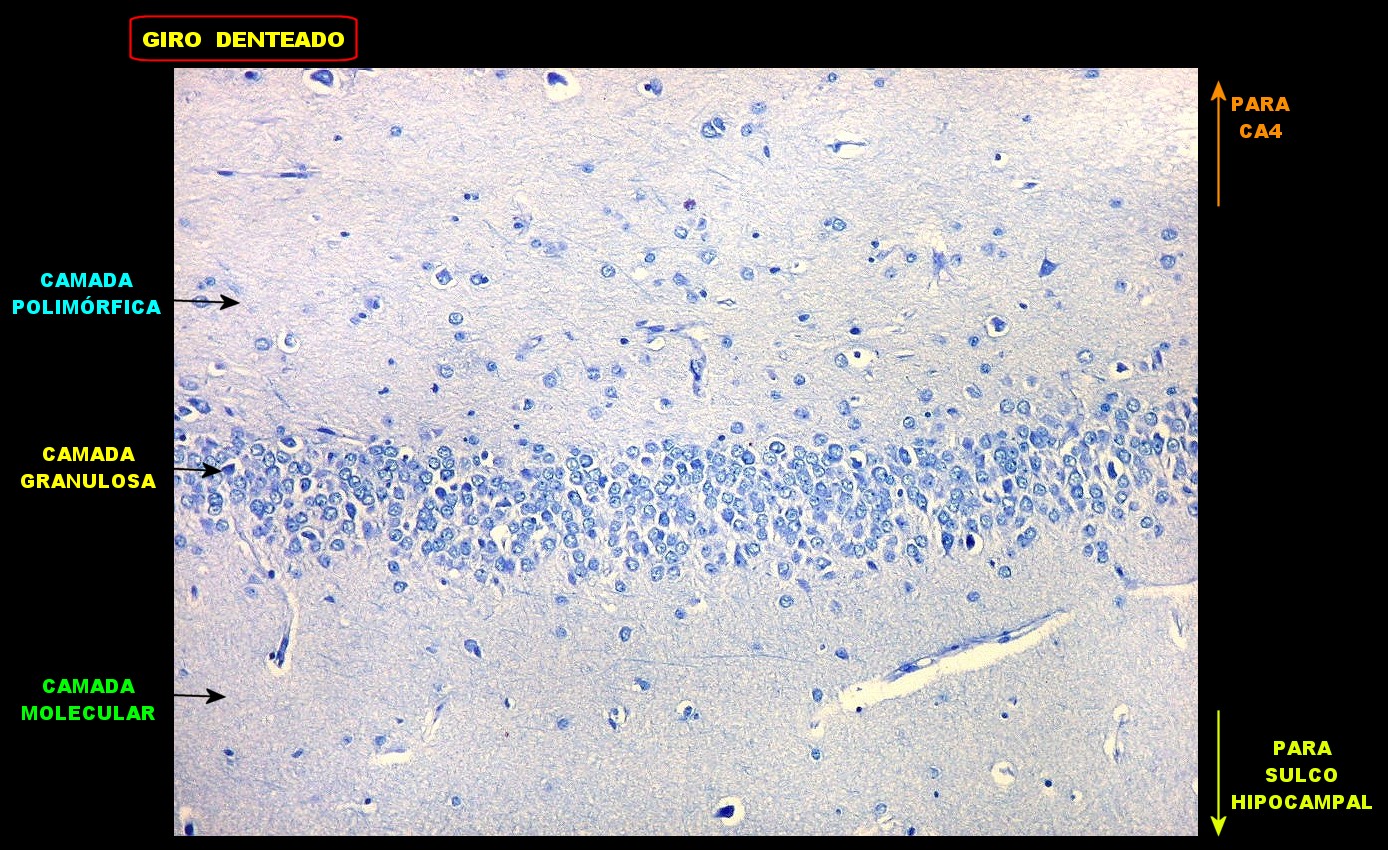
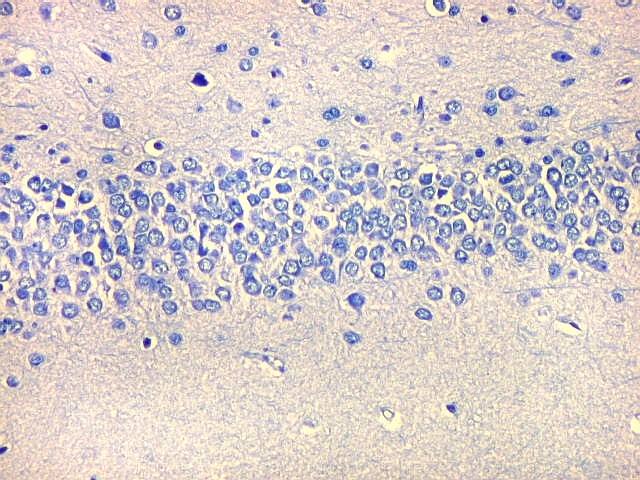
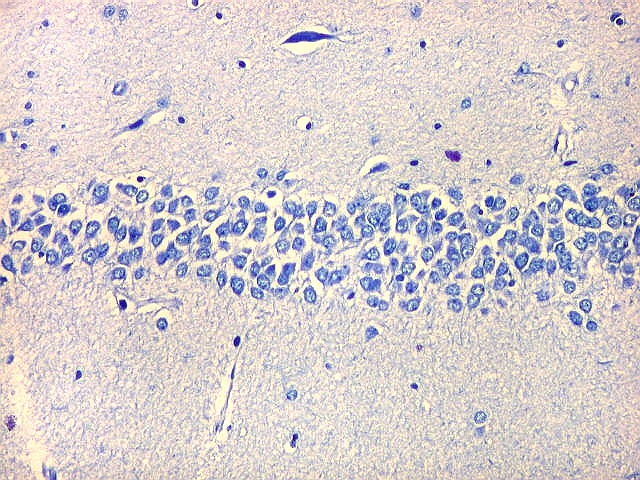
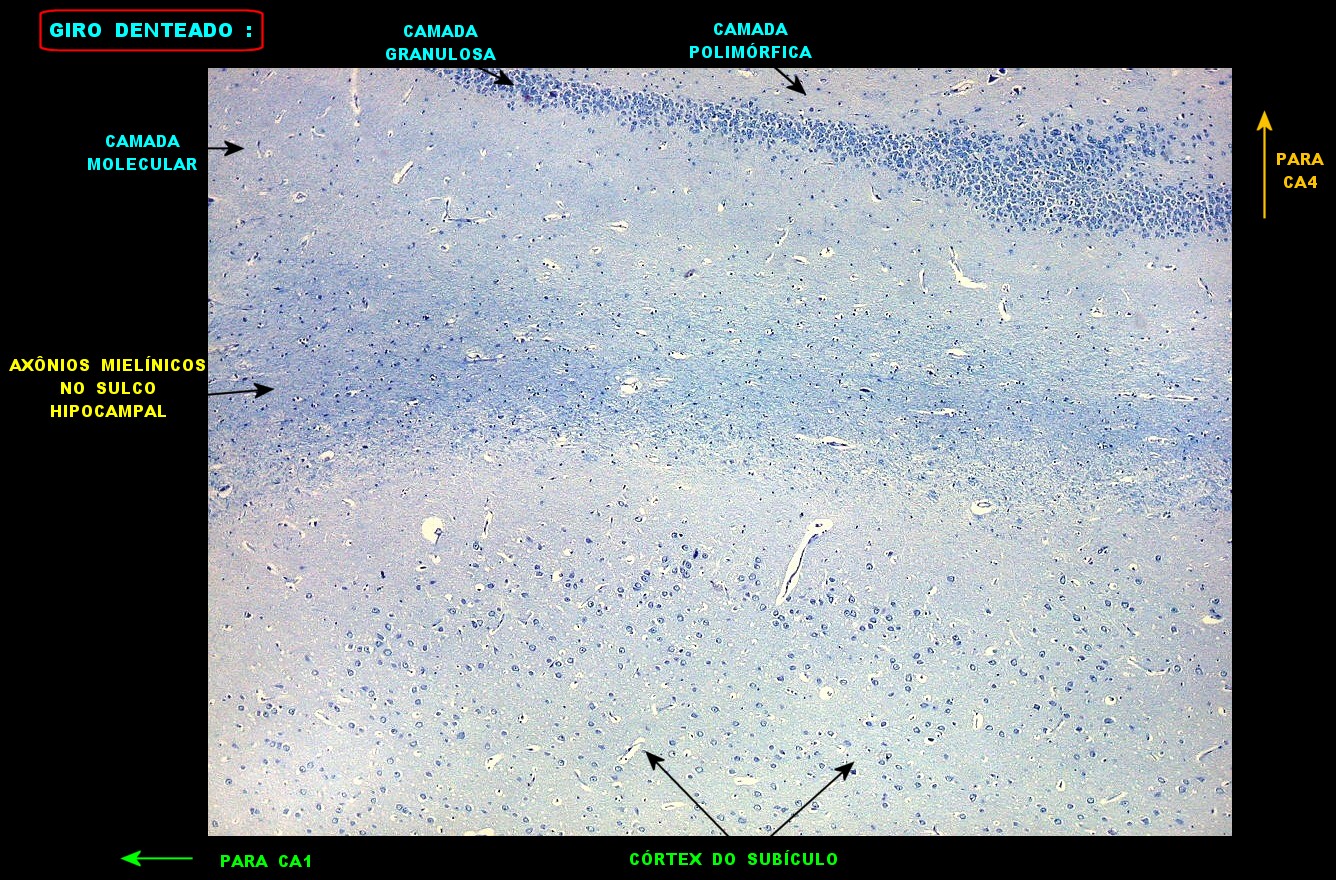
| Giro denteado. A camada granulosa, que contém os corpos dos neurônios granulosos, é a mais evidente do giro denteado. Logo acima vem a camada polimórfica, que faz transição para CA4. Abaixo, a camada molecular, que tem poucos corpos celulares de neurônios, mas recebe muitas fibras aferentes que fazem sinapse nos dendritos das células granulosas. Os axônios destas são as fibras musgosas que se distribuem a CA4 e CA3. Os neurônios granulosos têm corpo celular pequeno, com citoplasma escasso. O núcleo é grande, com nucléolo evidente. Para texto e esquema sobre as conexões do hipocampo, clique. |
 |
 |
 |
 |
 |
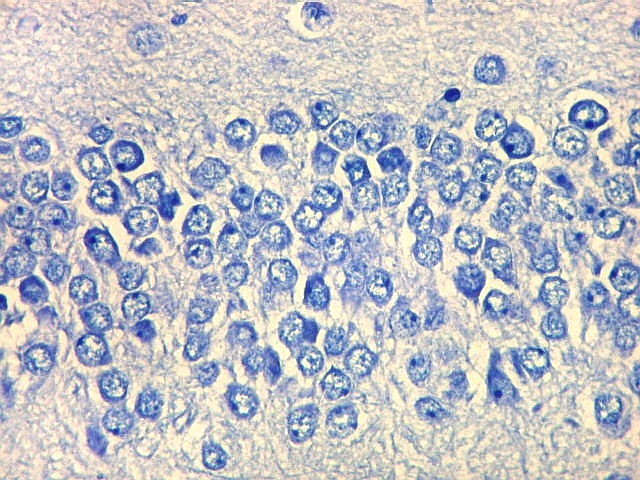 |
|
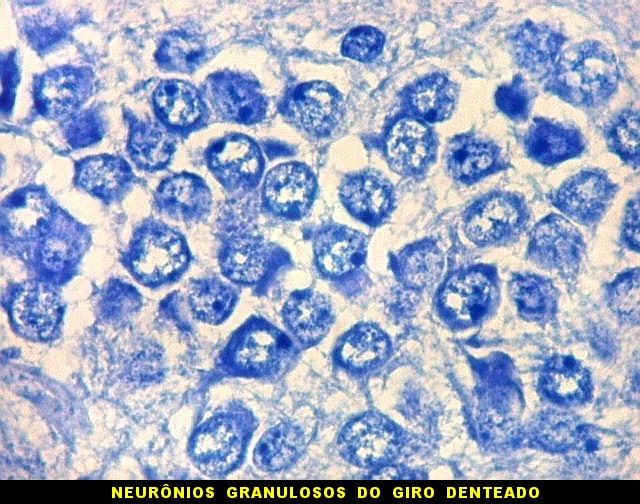 |
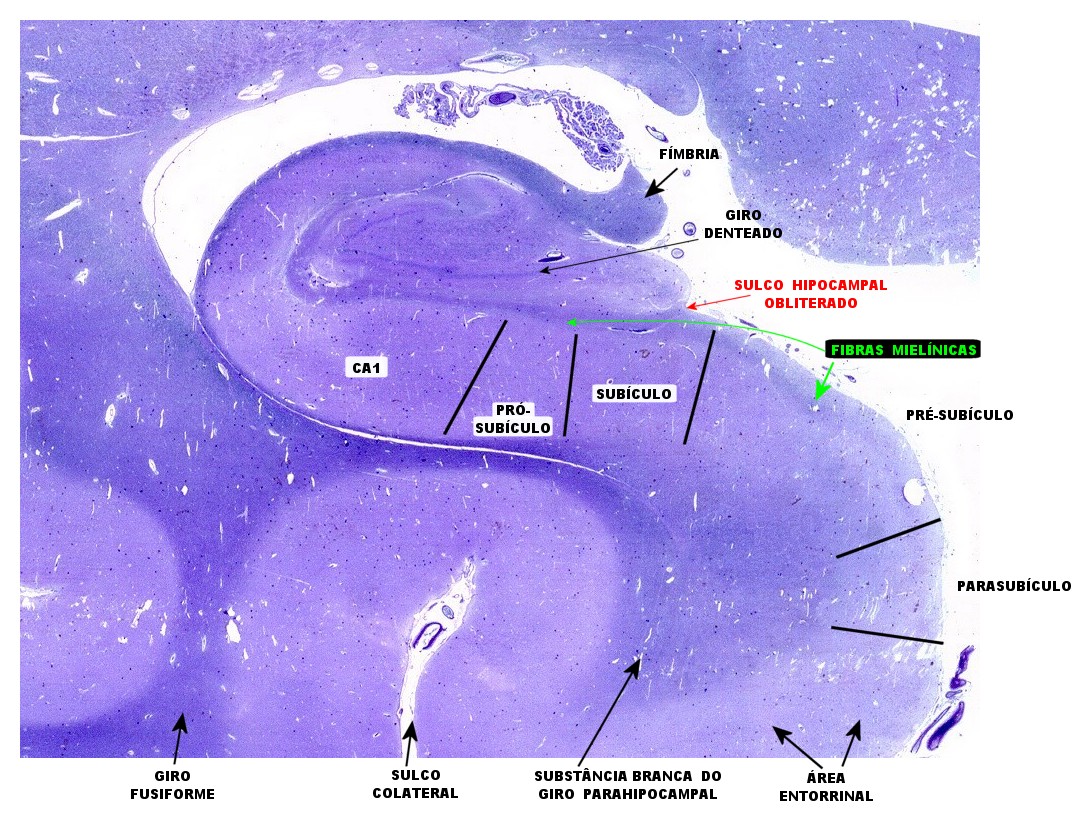
| Subículo
e divisões.
O subículo é a região do giro parahipocampal que faz transição entre a zona entorrinal na margem inferior do giro com o hipocampo acima. Reconhecem-se 4 sub-regiões, denominadas para-subículo, pré-subículo, subículo propriamente dito e pró-subículo, indo da zona entorrinal para o hipocampo. Para detalhes, clique. |
 |
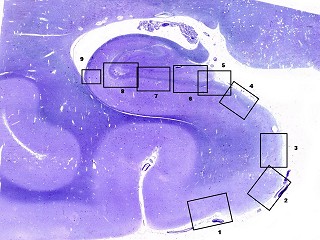
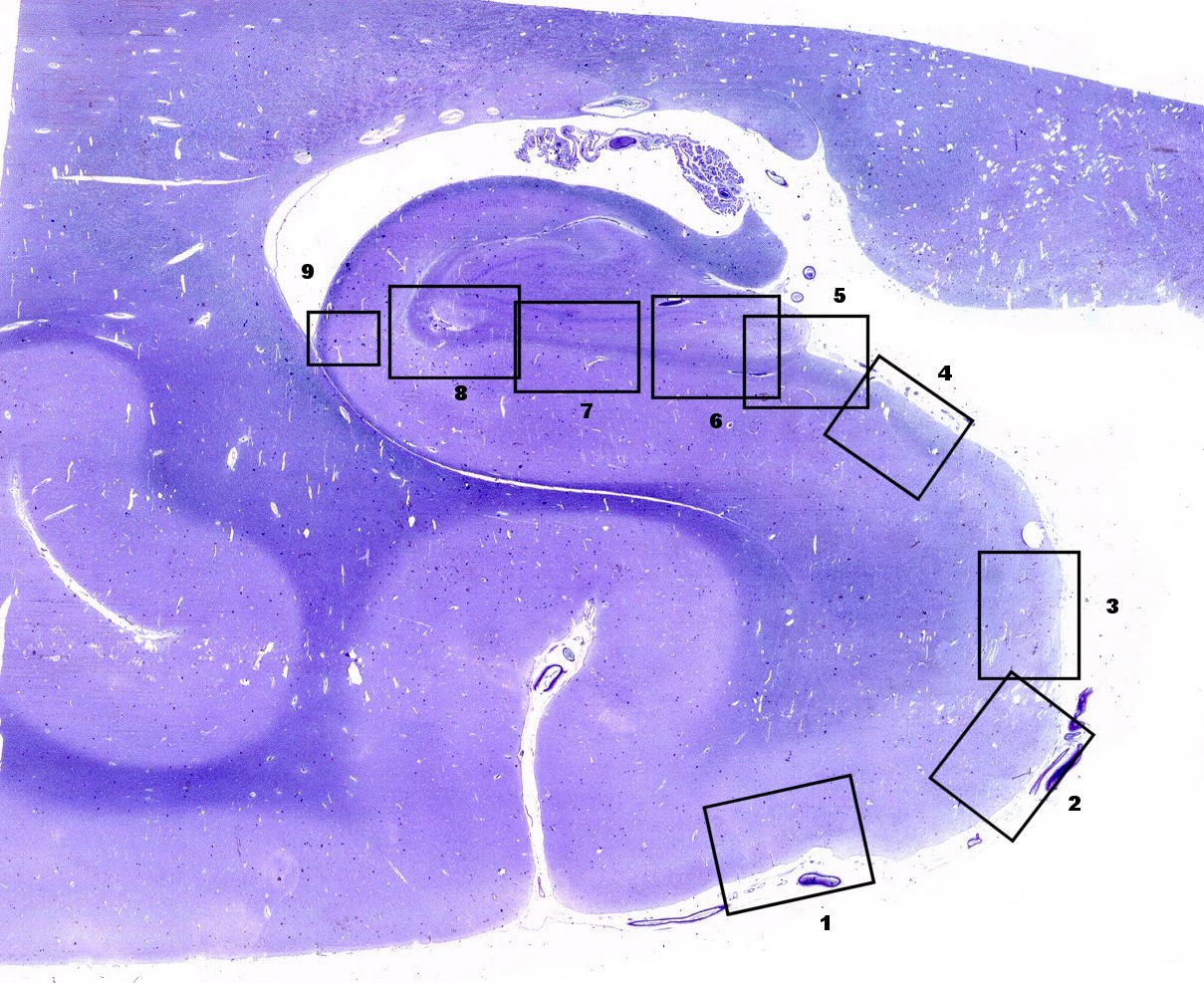
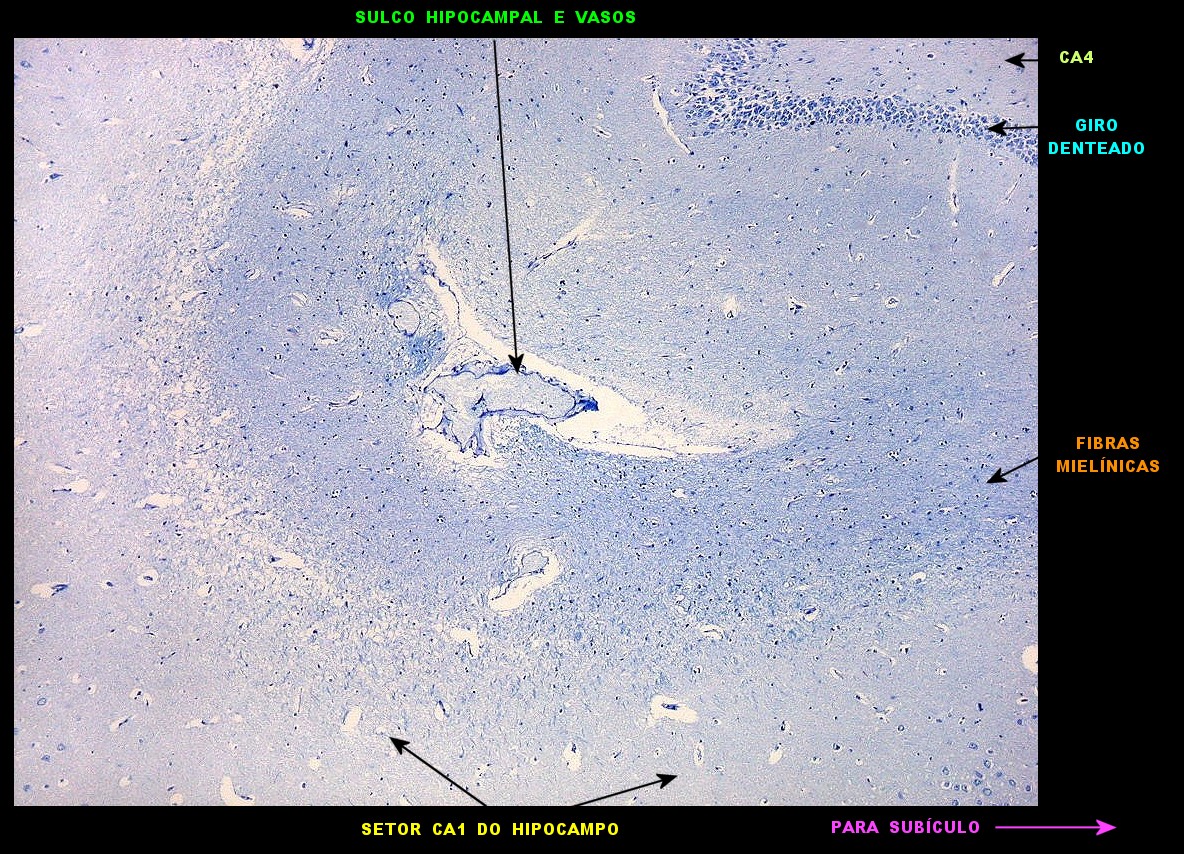
| Seqüência
pela zona entorrinal, subículo e divisões, sulco hipocampal
e CA1. A lâmina
escaneada com 1200 dpi abaixo serve de mapa para as fotos pela superfície
da zona entorrinal (1, 2), parasubículo (3), pré-subículo
(4), sulco hipocampal (5 a 8) obliterado, onde se observam fibras mielínicas
em continuação com as da camada mais superficial do pré-subículo.
Em 8 há um vaso remanescente no sulco hipocampal obliterado. Clique
nos retângulos para ir diretamente aos detalhes (fotos no microscópio
com objetiva 4x).
Obs. as divisões foram baseadas nos esquemas do livro de Duvernoy, 1988. Não são precisas e servem apenas como referência. Por exemplo, a própria existência do pró-subículo é questionada por alguns autores. |
 |
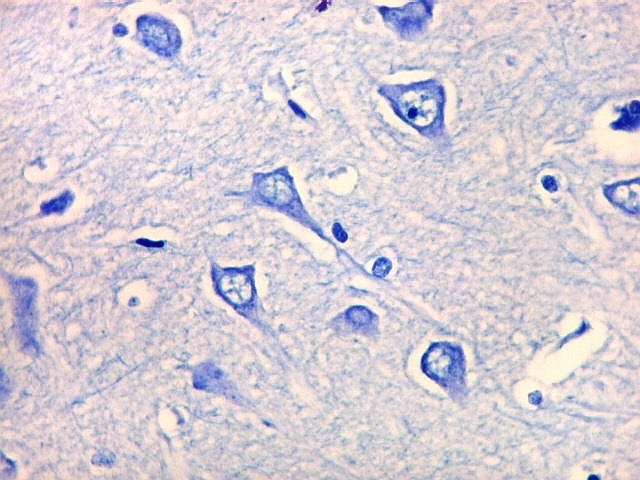
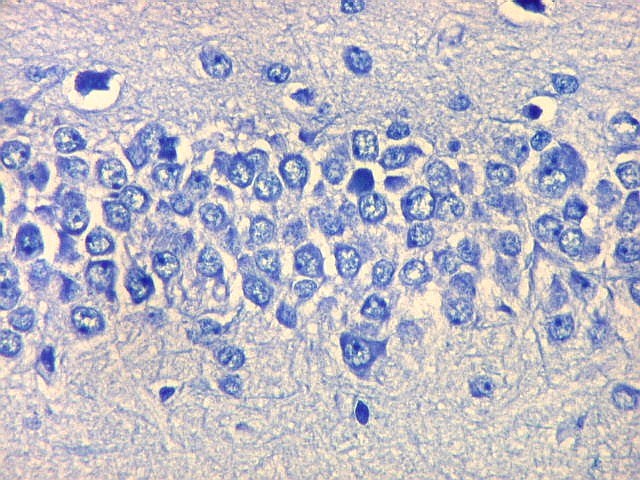
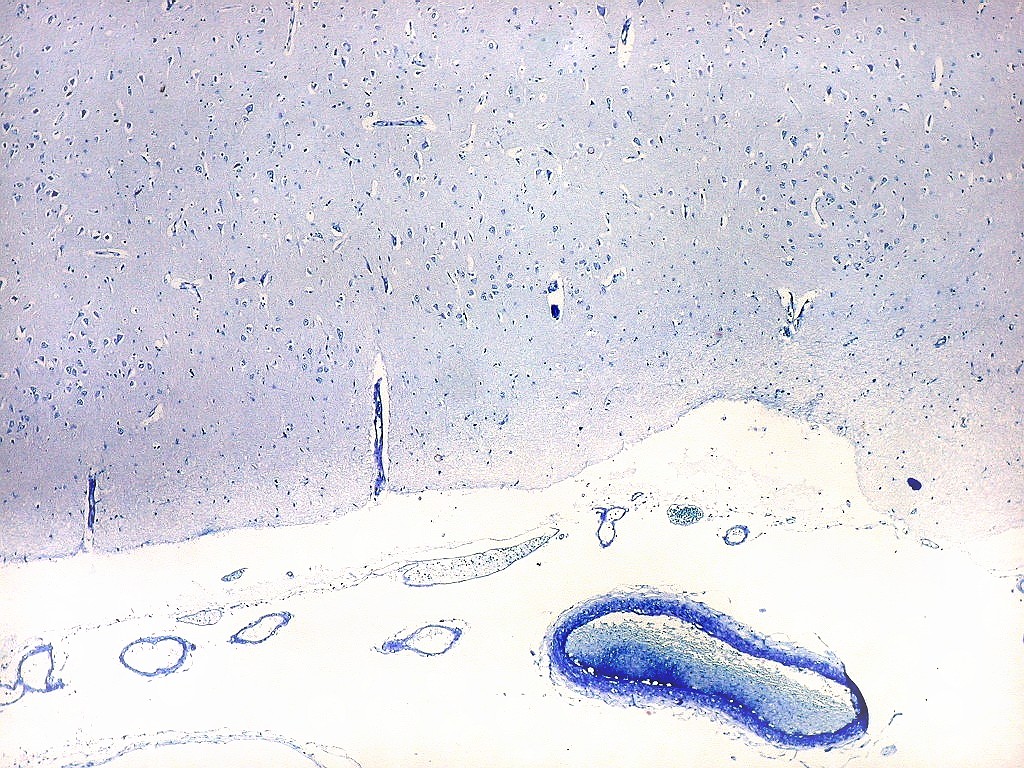
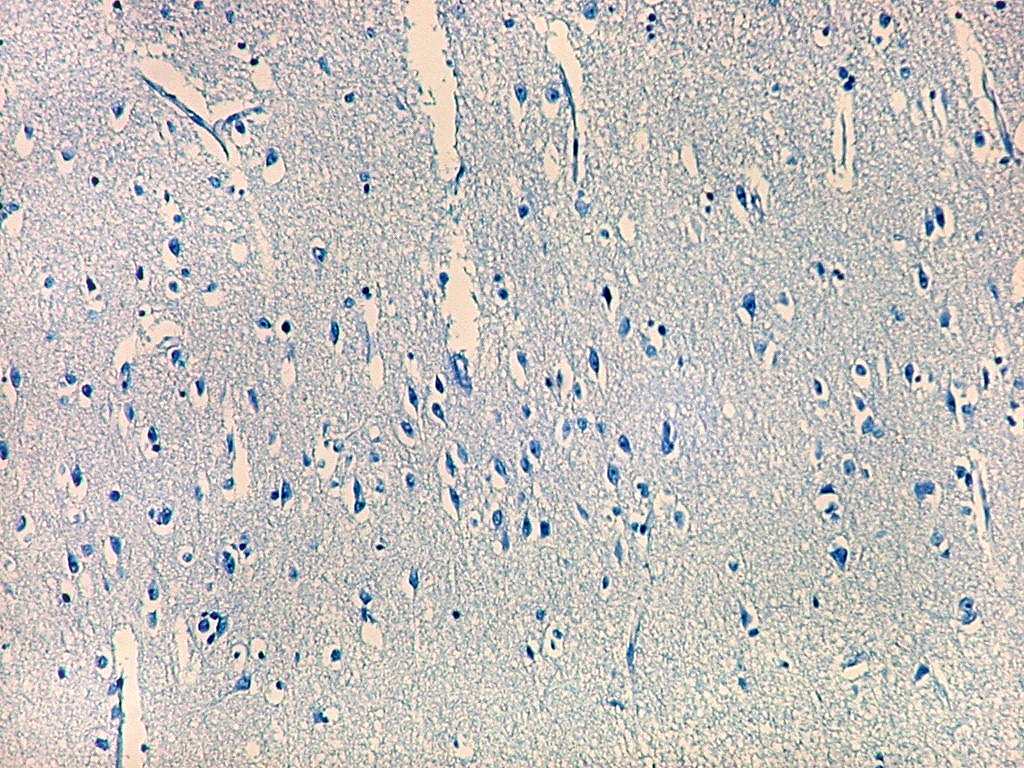
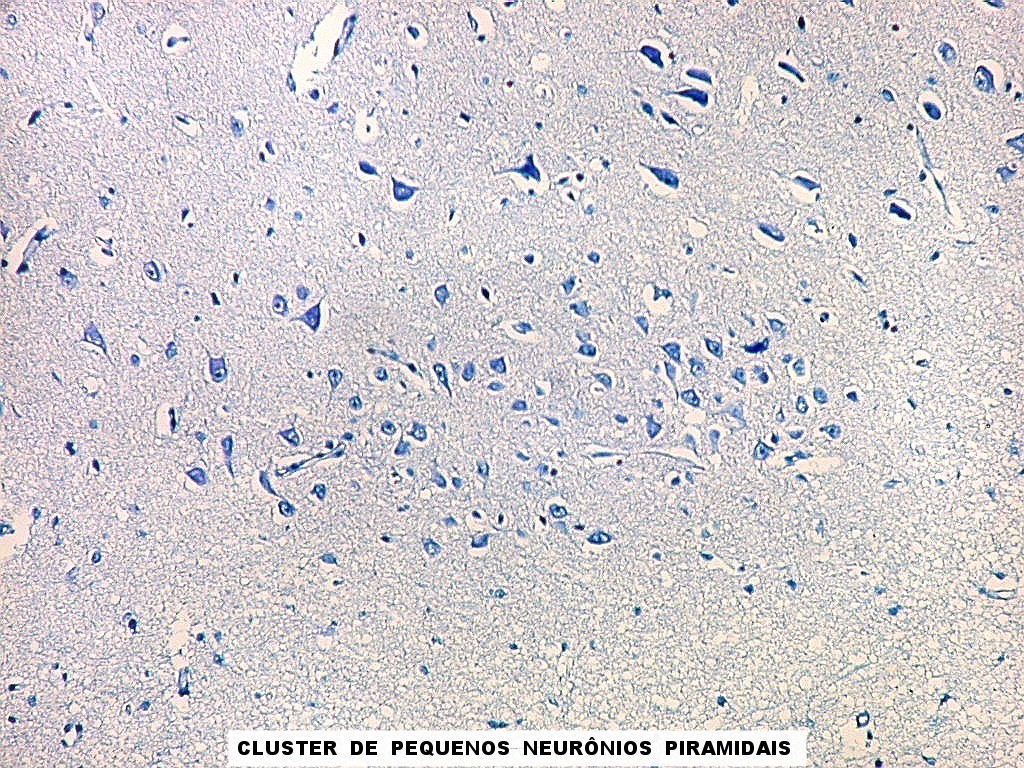
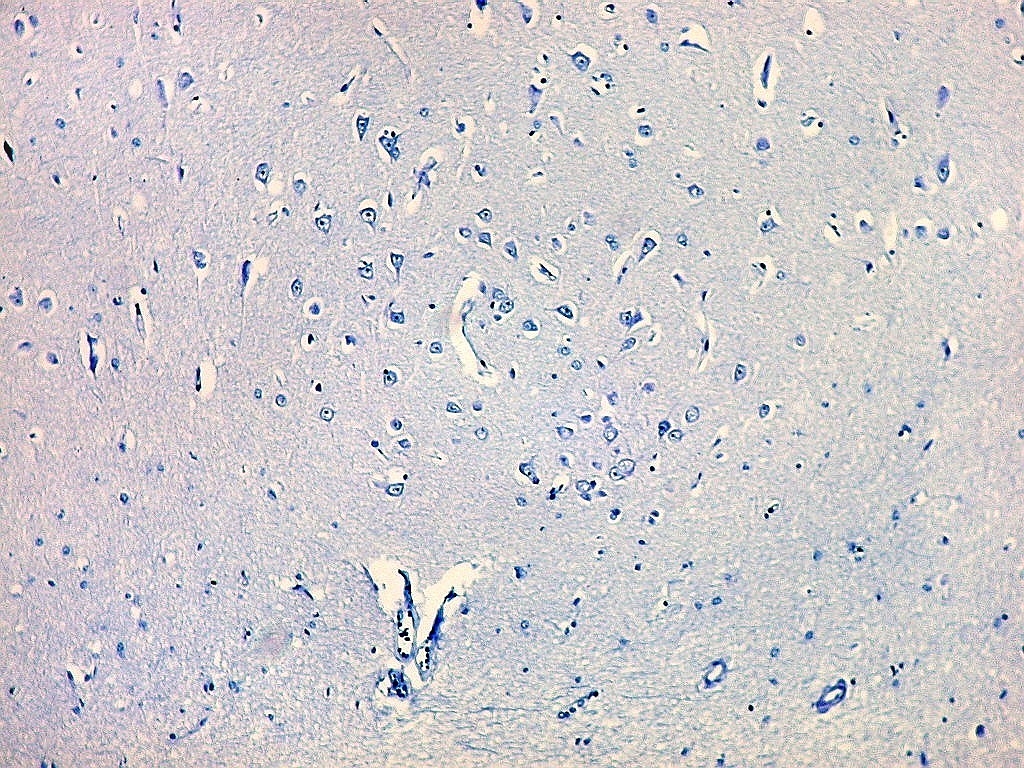
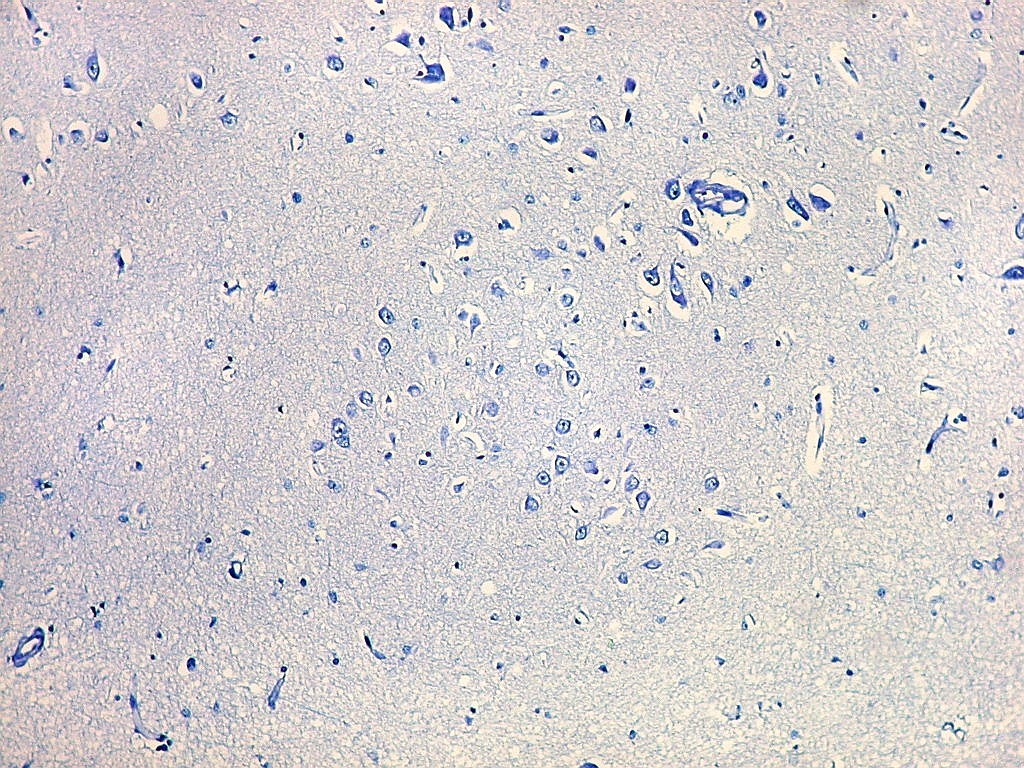
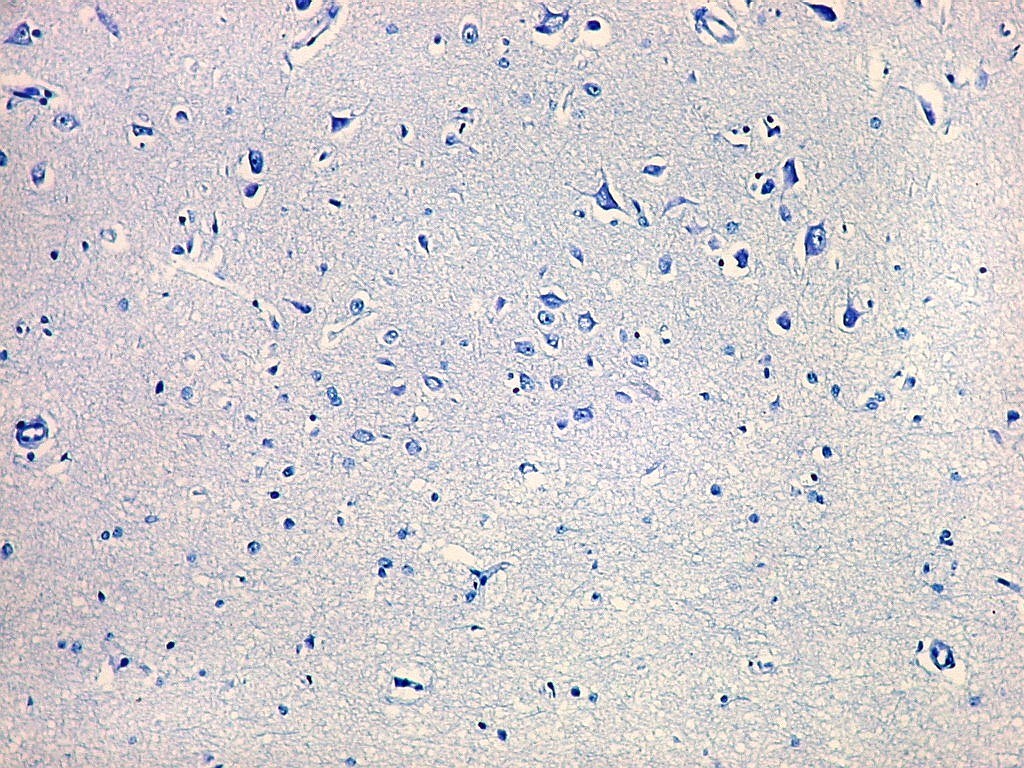
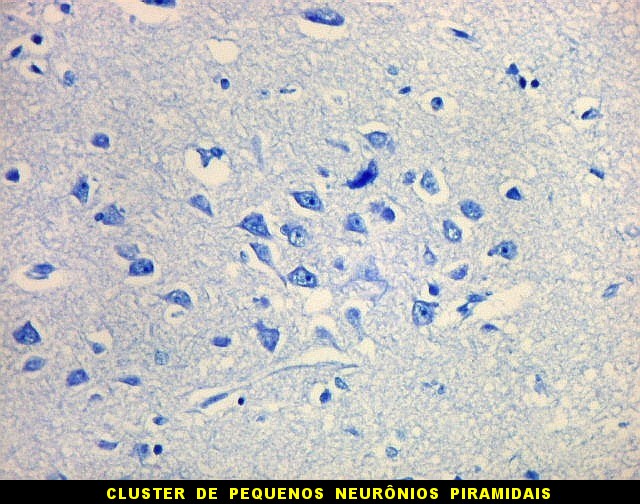
| Campo 1 - zona entorrinal. As fotos abaixo foram tomadas na camada superficial da face medial do giro parahipocampal, indo da área entorrinal em direção ao hipocampo. Observam-se na segunda camada do córtex agrupamentos de pequenos neurônios piramidais formando 'clusters', com áreas pobres em corpos celulares entre os aglomerados. Embora na literatura seja descrito que tais aglomerados ocorrem mais notavelmente no pré-subículo, encontramos os mesmos desde a área entorrinal neste preparado. Esta arquitetura é característica da face medial do giro parahipocampal e torna sua identificação histológica relativamente fácil. |
 |
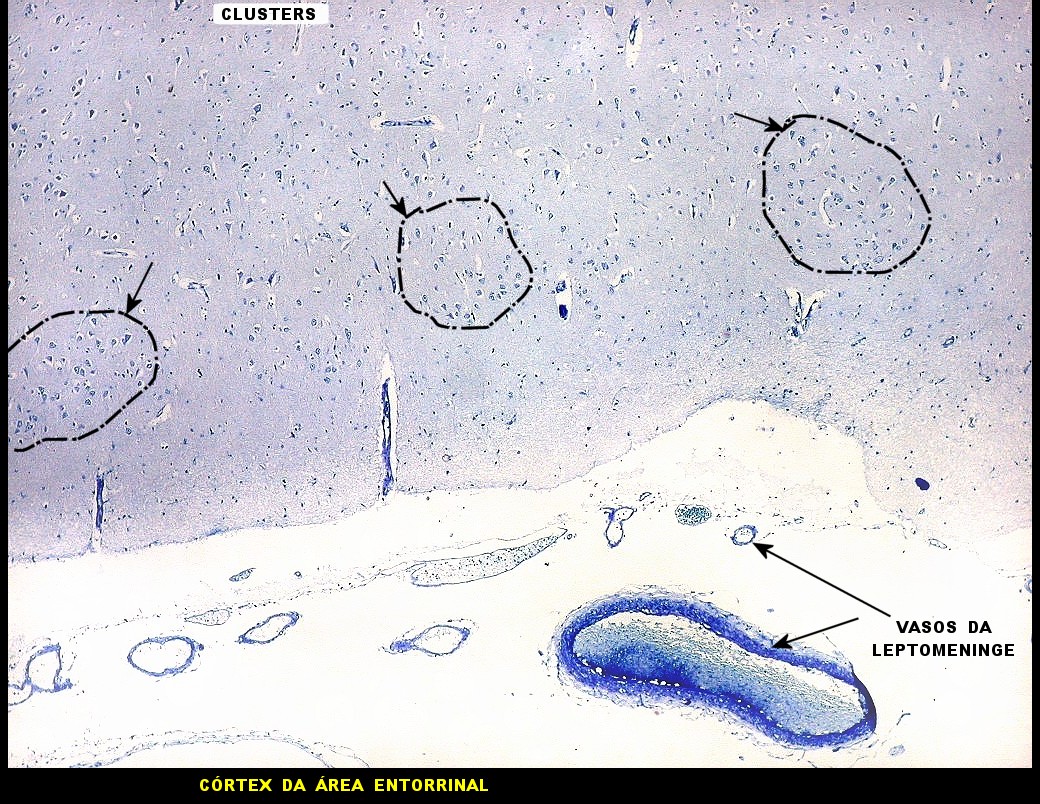 |
| Campo 2 - zona entorrinal. Fotos 1 e 2 com leptomeninge para baixo (=posição anatômica). |
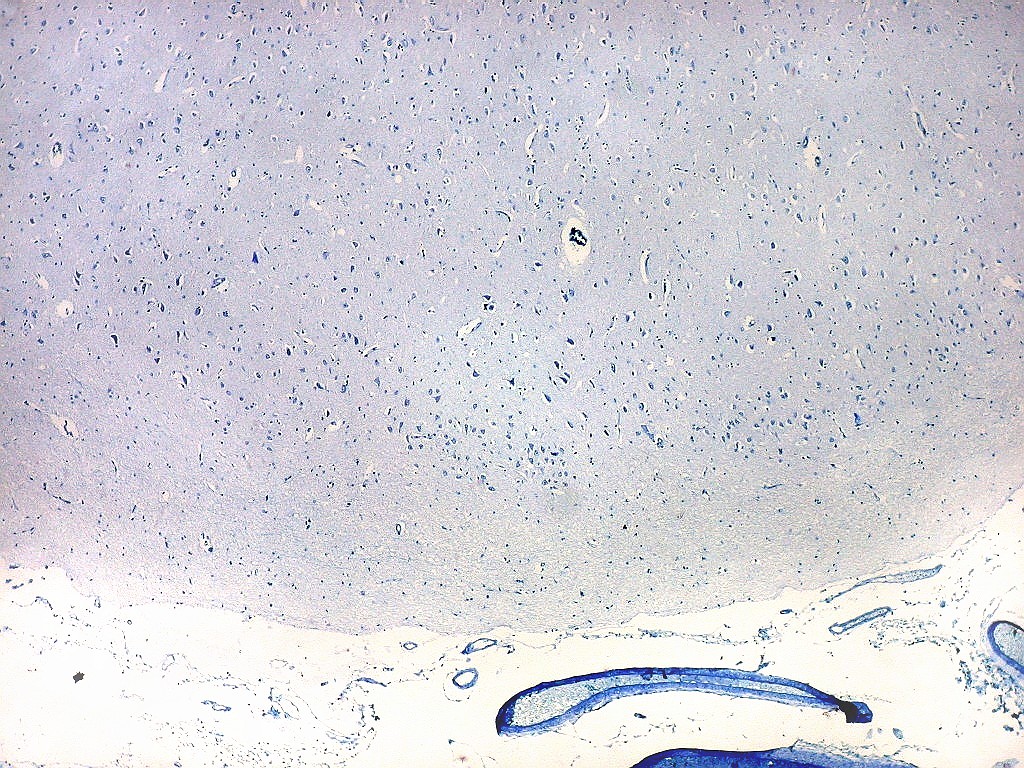 |
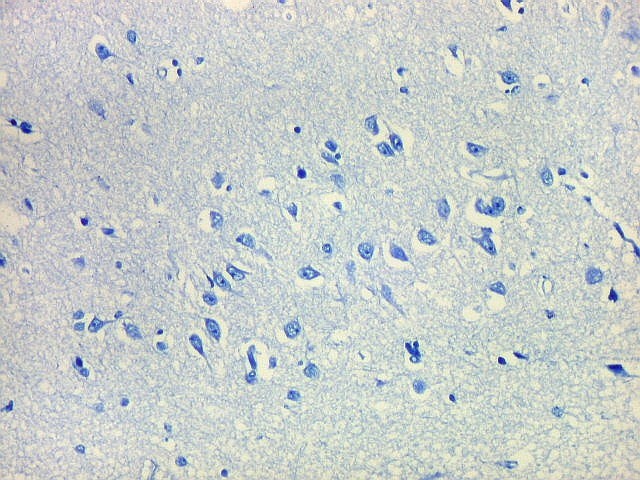
| Campos 1 e 2 - Fotos com objetiva 10x mostrando os clusters de pequenos neurônios piramidais na camada superficial do córtex do giro parahipocampal. |
 |
 |
 |
 |
 |
 |
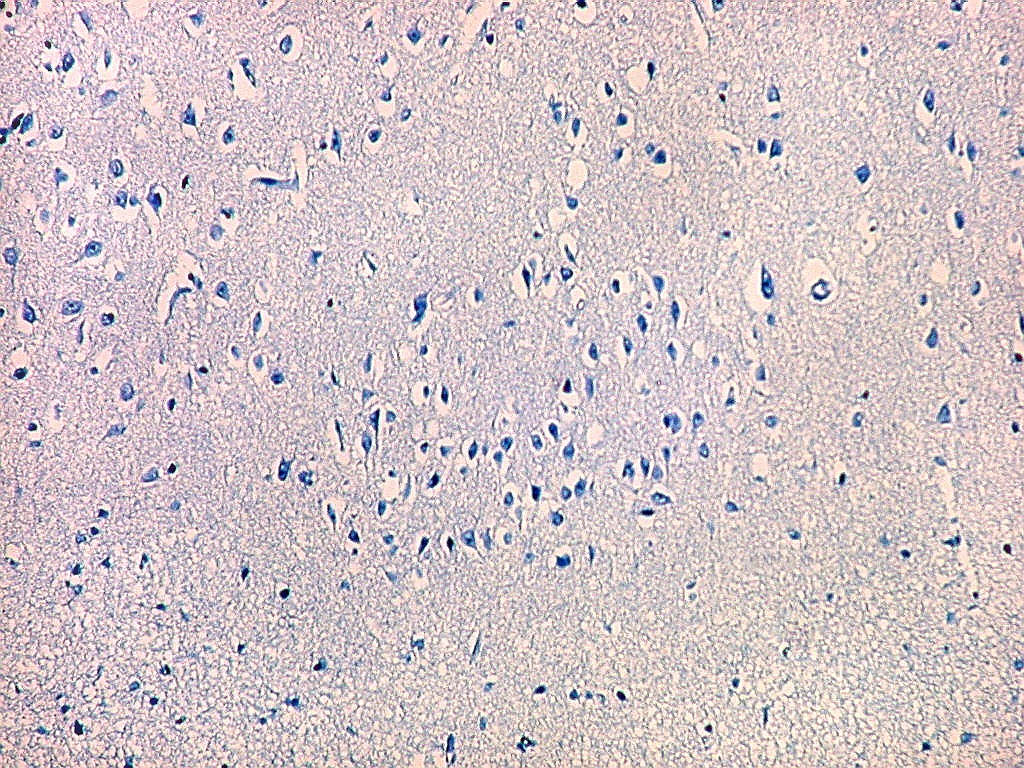
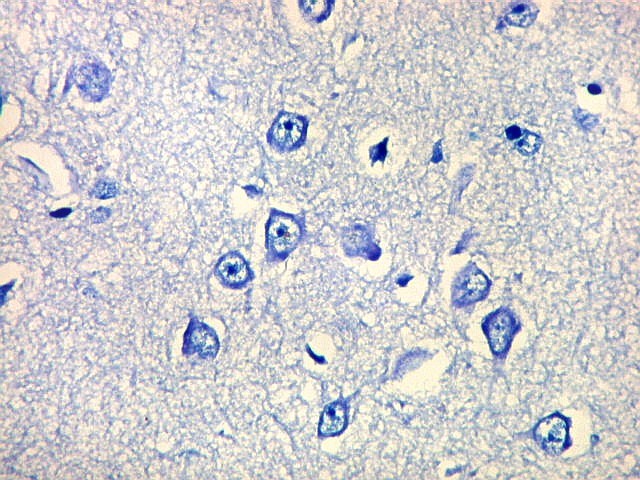
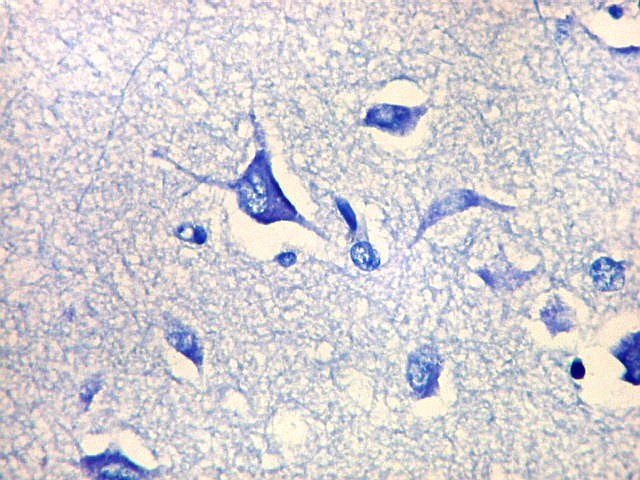
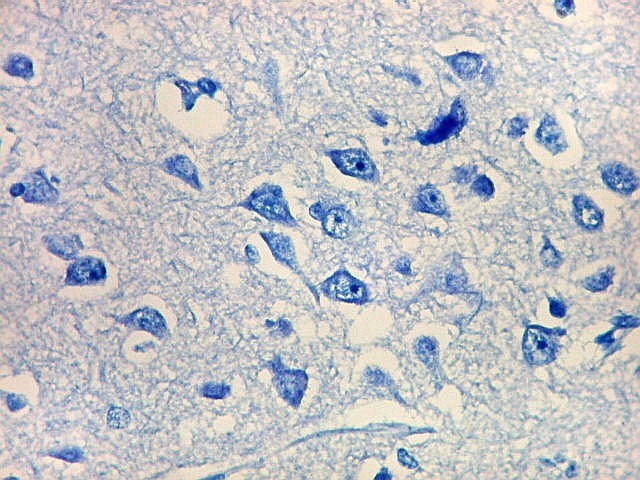
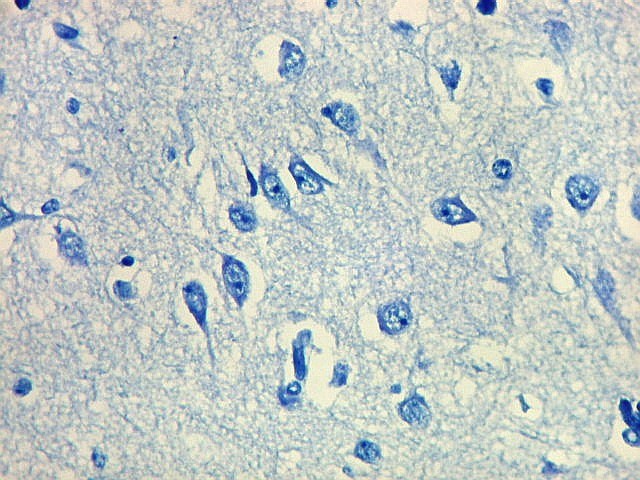
| Detalhes, objetivas 20x e 40x. Aqui percebe-se que os neurônios dos clusters são piramidais pequenos, com dendrito apical voltado para a leptomeninge (para baixo nestas fotos). | |
 |
|
 |
 |
 |
 |
 |
 |
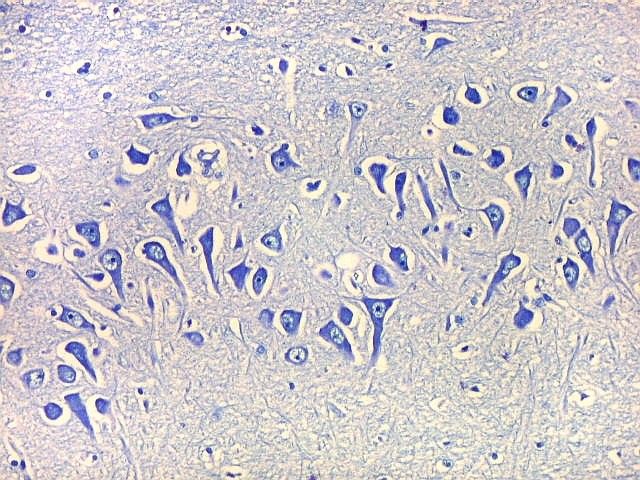
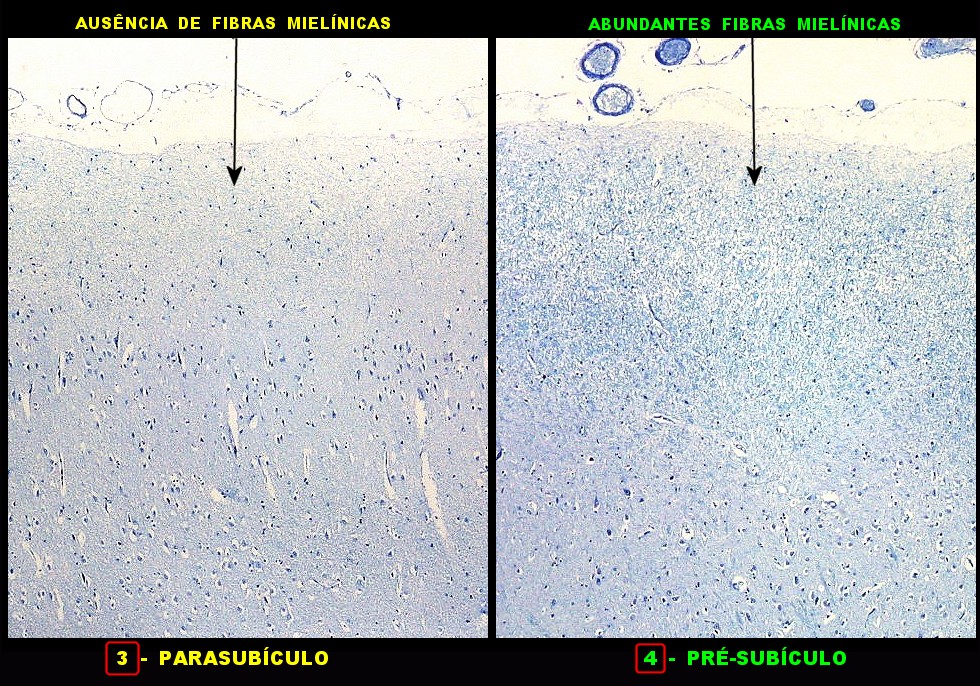
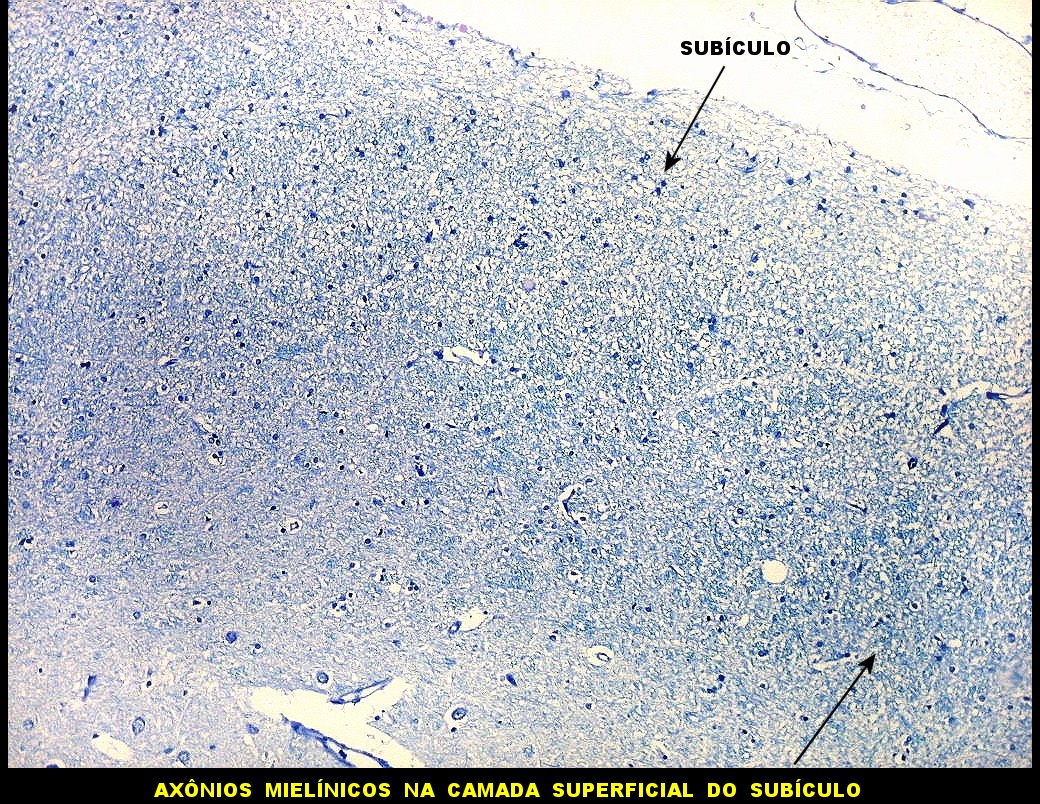
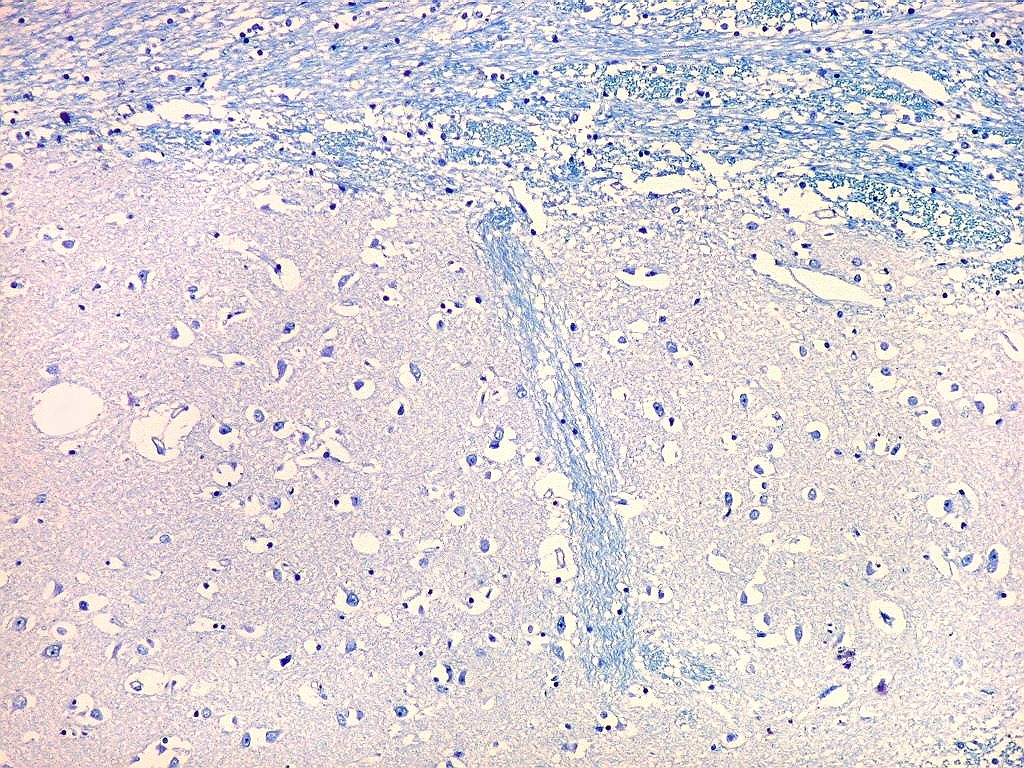
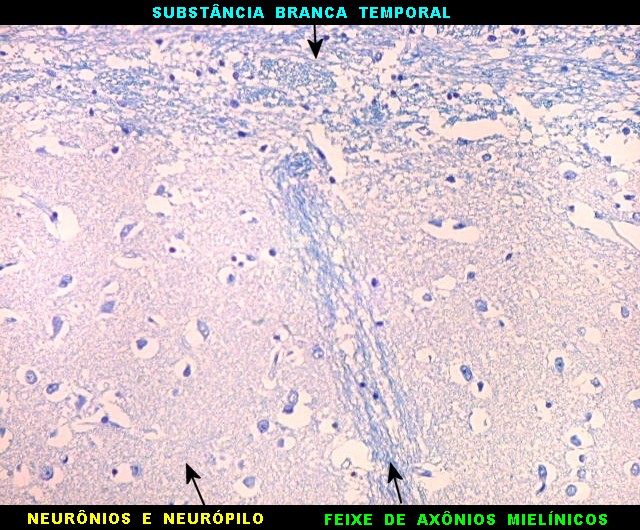
| Fibras mielínicas na camada molecular do giro parahipocampal. As fotos seguintes foram tomadas acompanhando a camada mais superficial do córtex do giro parahipocampal na região do parasubículo e pré-subículo. Nas regiões mais distantes do hipocampo a camada molecular não tem fibras mielínicas. À medida que se aproxima do hipocampo, aparece um contingente de fibras mielínicas que é notado pela cor azul turquesa no LFB-Nissl. Estas fibras continuam pelo sulco hipocampal. Aqui, as fotos acompanham a curvatura do giro parahipocampal, com leptomeninge para cima. Aspecto semelhante é observado nas preparações para neurofilamento, que demonstram axônios. |
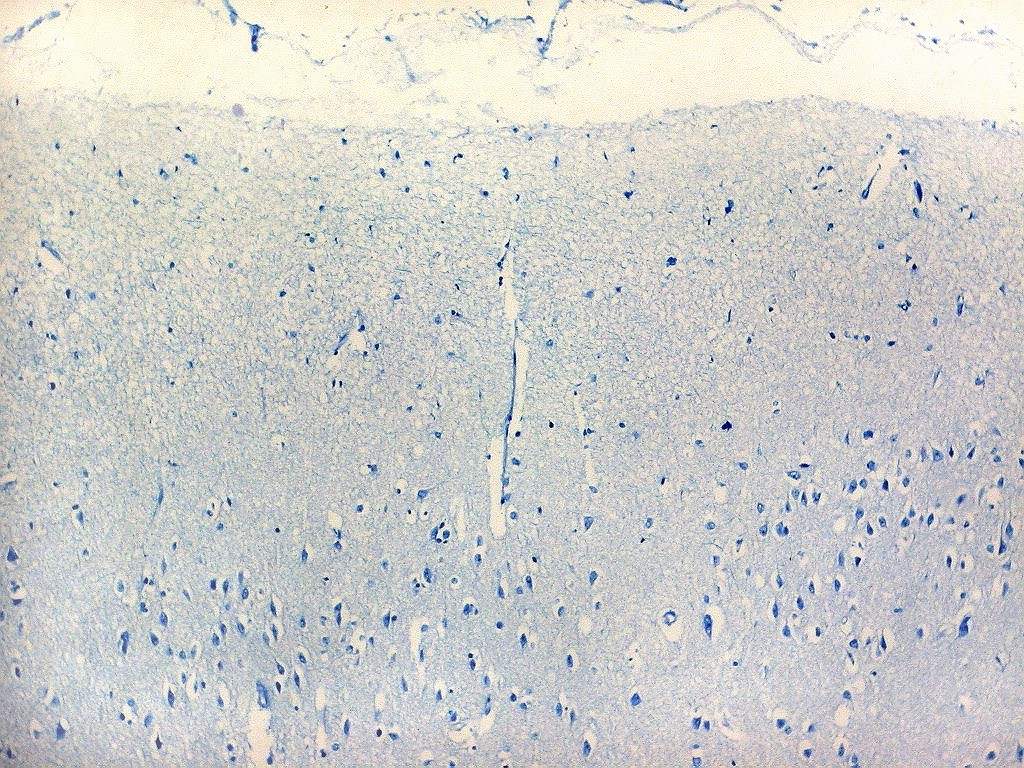
| Campo 3, parasubículo. |
 |
 |
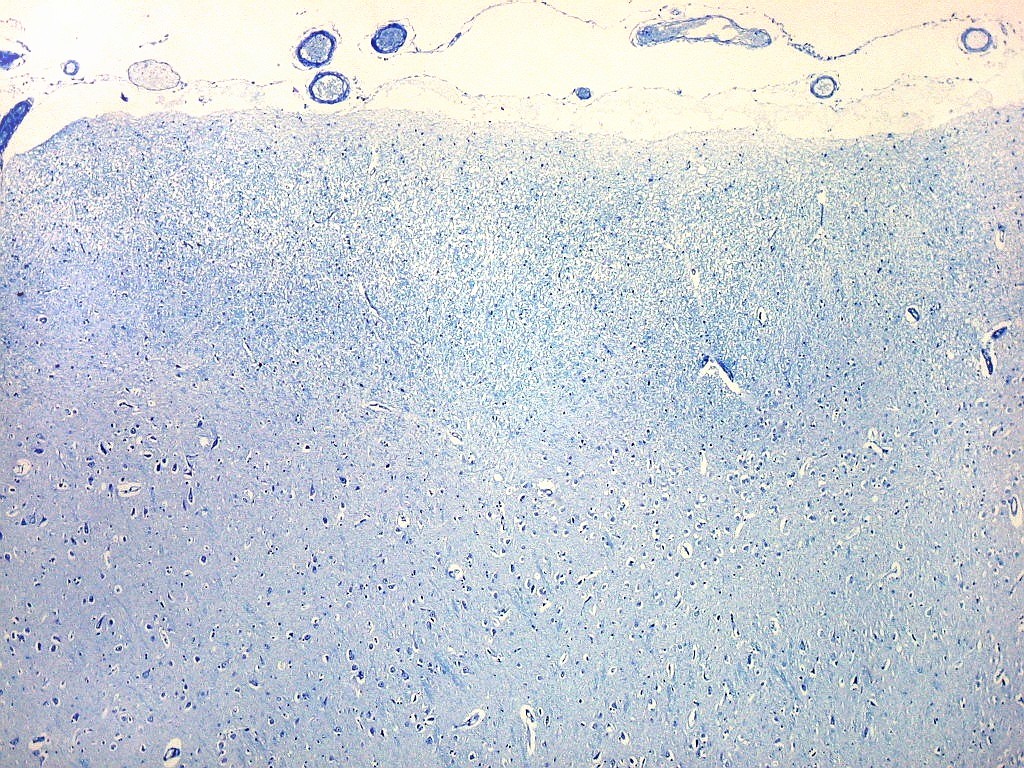
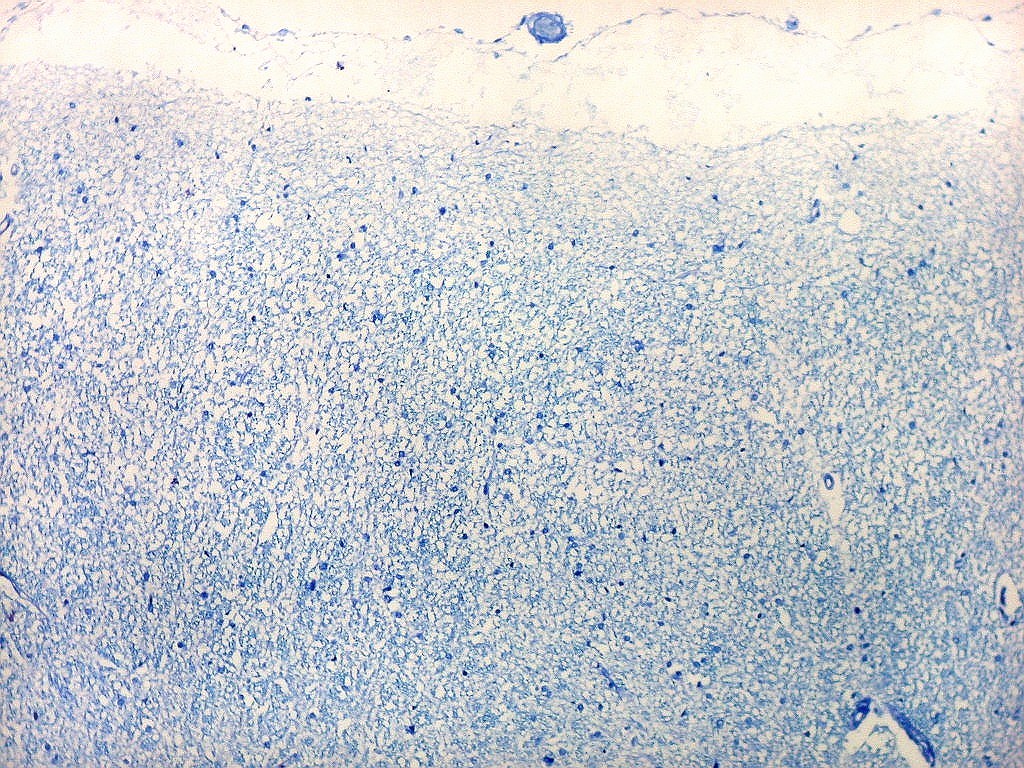
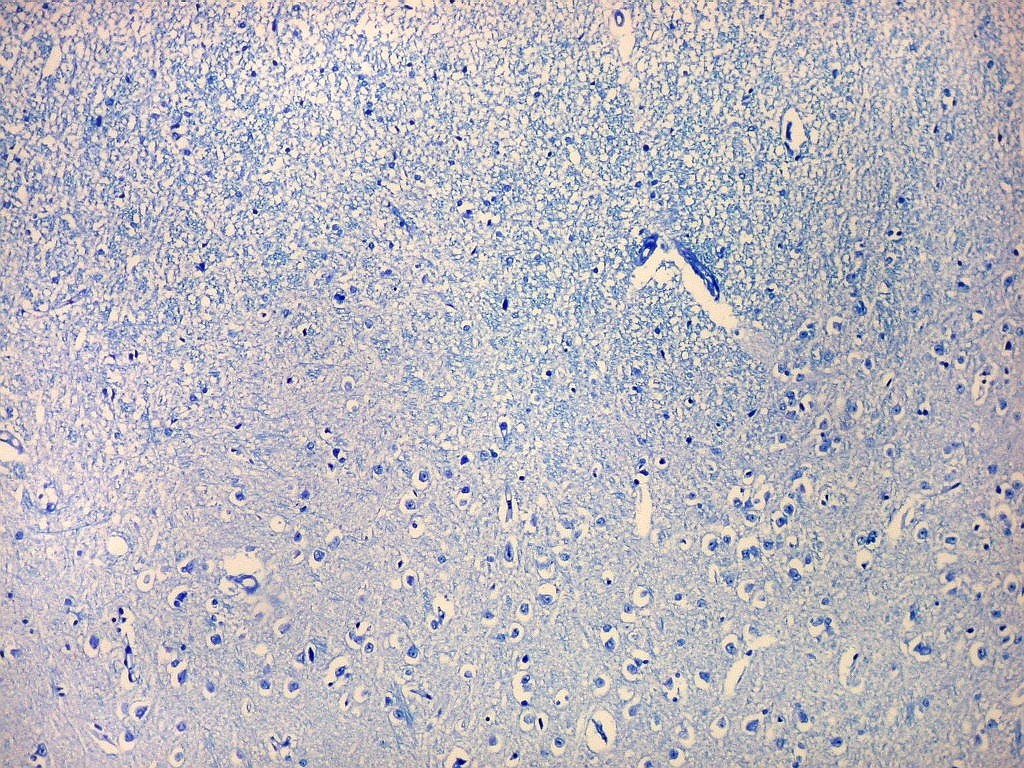
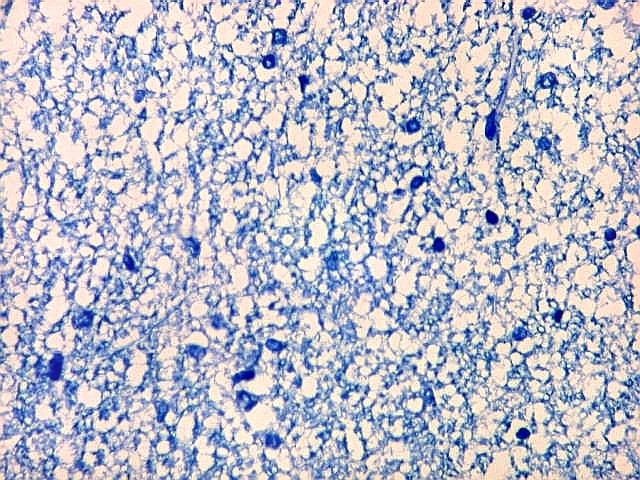
| Campo 4, pré-subículo. Neste nível notam-se abundantes fibras mielínicas na camada mais superficial do córtex parahipocampal. Admitimos que se trate de fibras aferentes que vão constituir a via perfurante, que segue o sulco hipocampal nas fotos seguintes. Para texto sobre anatomia do hipocampo e texto e esquema das conexões, clique. |
 |
 |
 |
 |
 |
 |
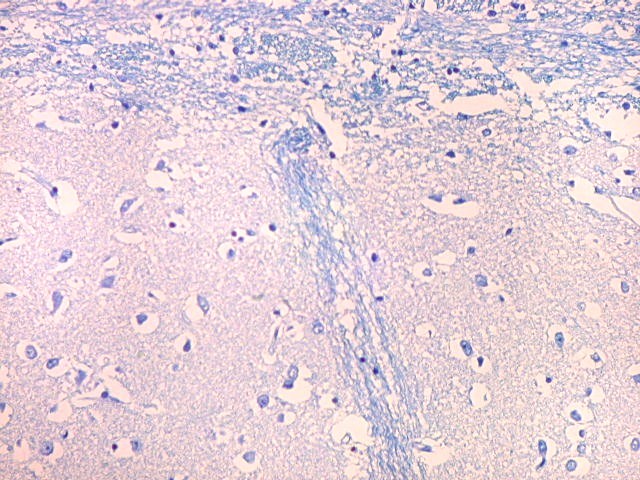
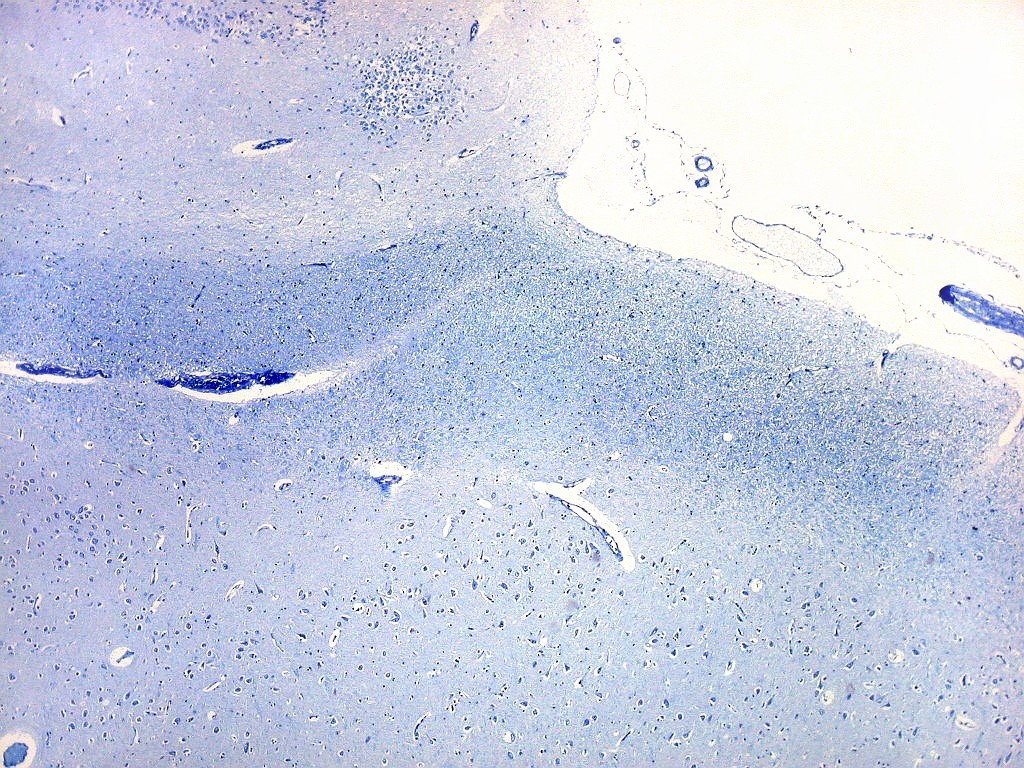
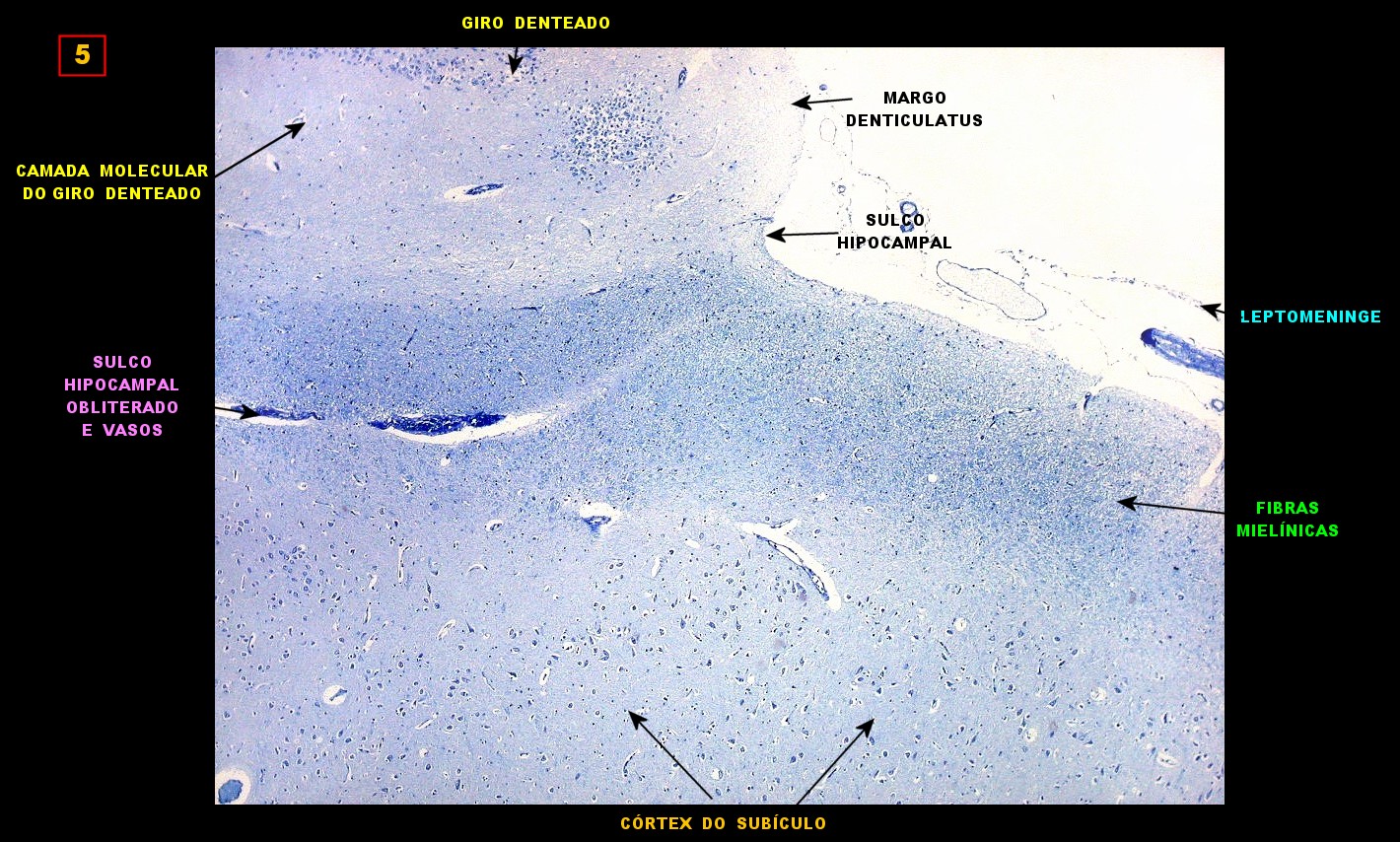
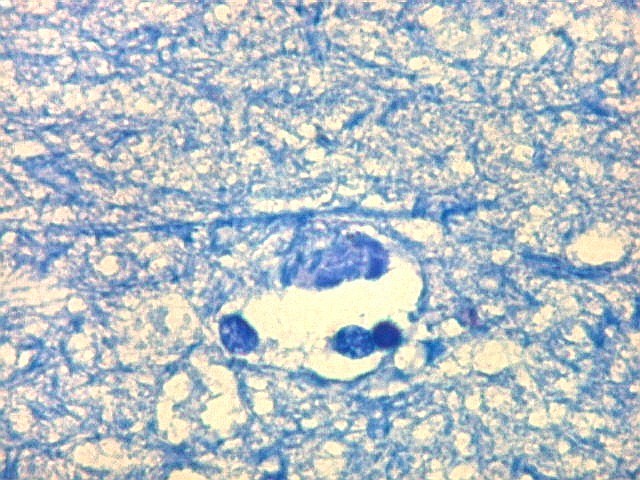
| Campo 5, sulco hipocampal, extremidade medial. Ao longo do sulco hipocampal obliterado observam-se numerosas fibras mielínicas que, segundo os esquemas de conexões do hipocampo, provêm da área entorrinal e distribuem aferências ao giro denteado (cima) e ao córtex hipocampal abaixo. Há também vasos que originalmente se localizavam na leptomeninge entre o hipocampo e o giro denteado. Com a fusão das duas estruturas durante a vida fetal, os vasos ficam embutidos no próprio tecido nervoso. |
 |
 |
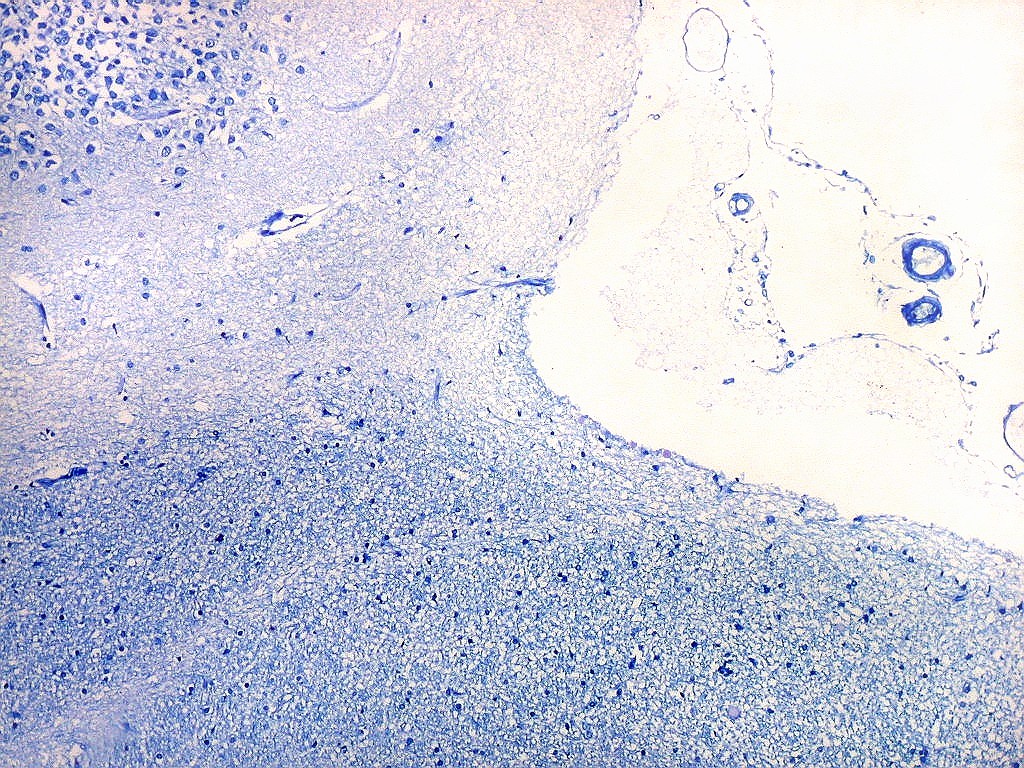 |
 |
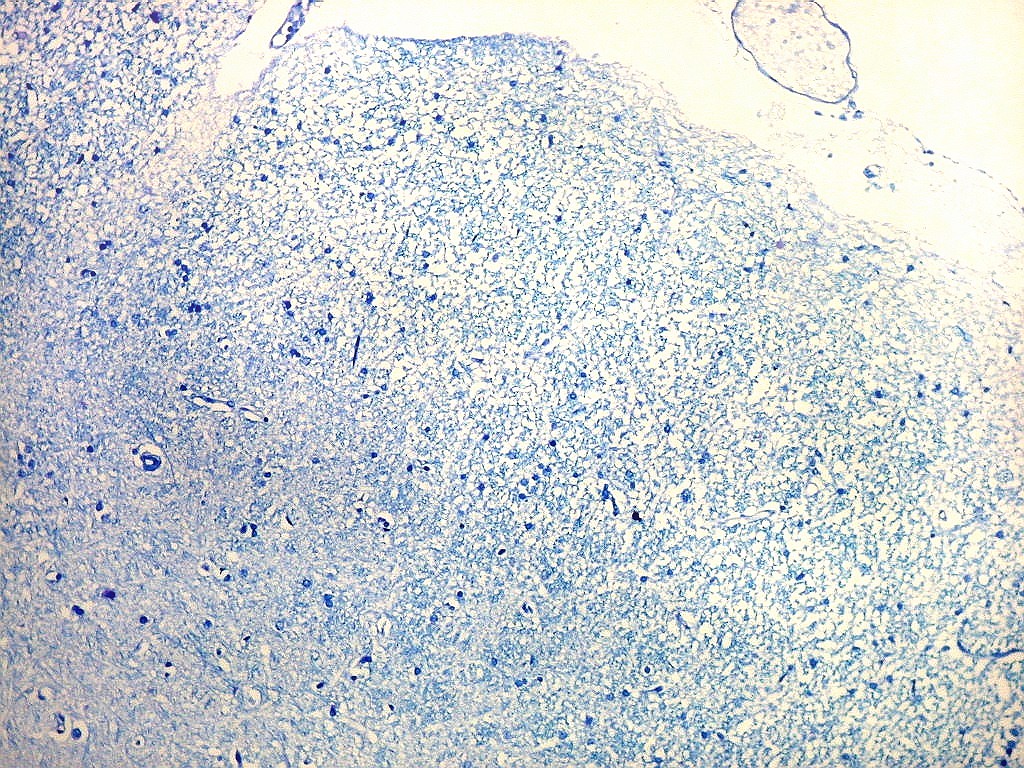 |
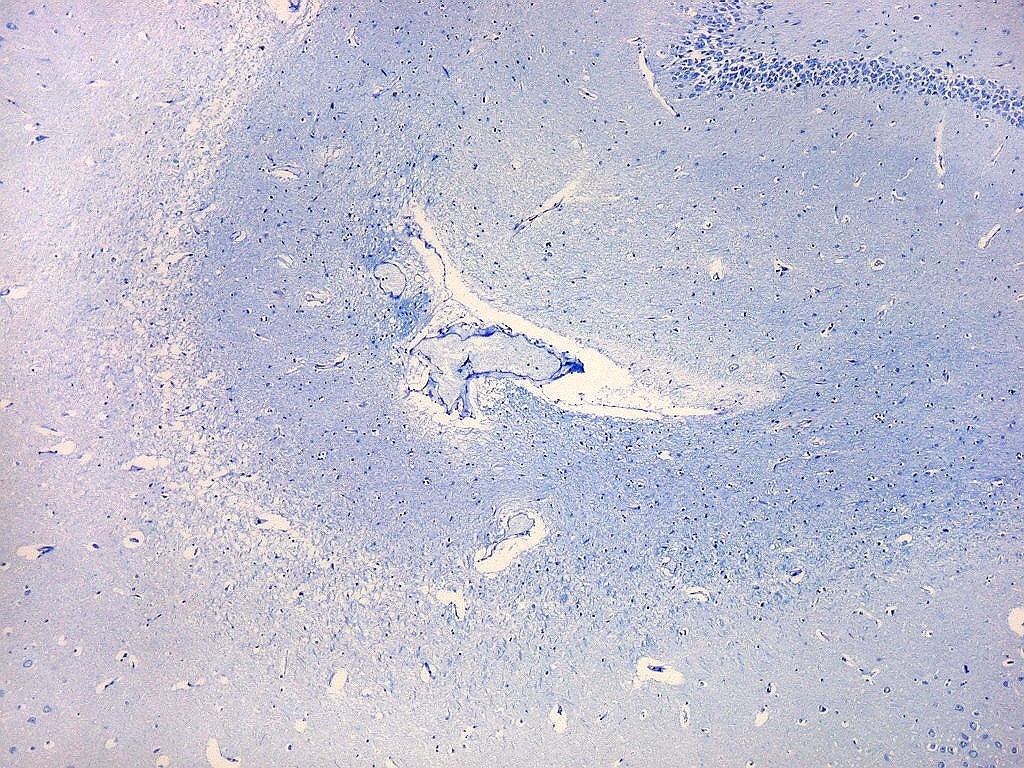
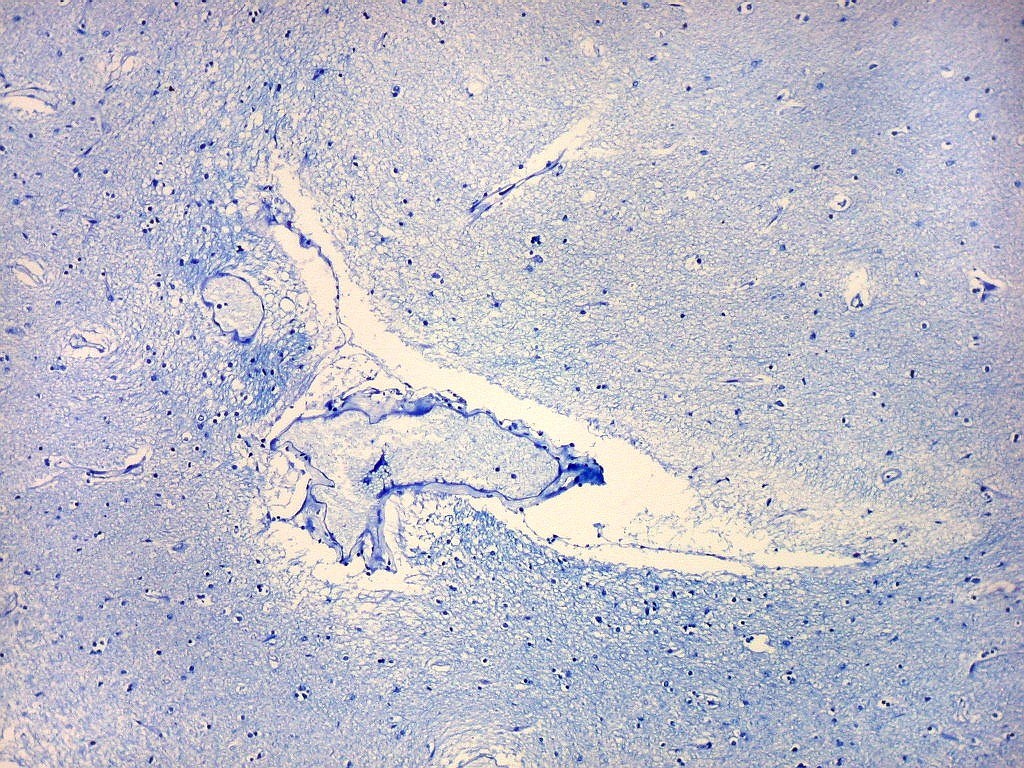
| Campo 6, sulco hipocampal, entre giro denteado e subículo |
 |
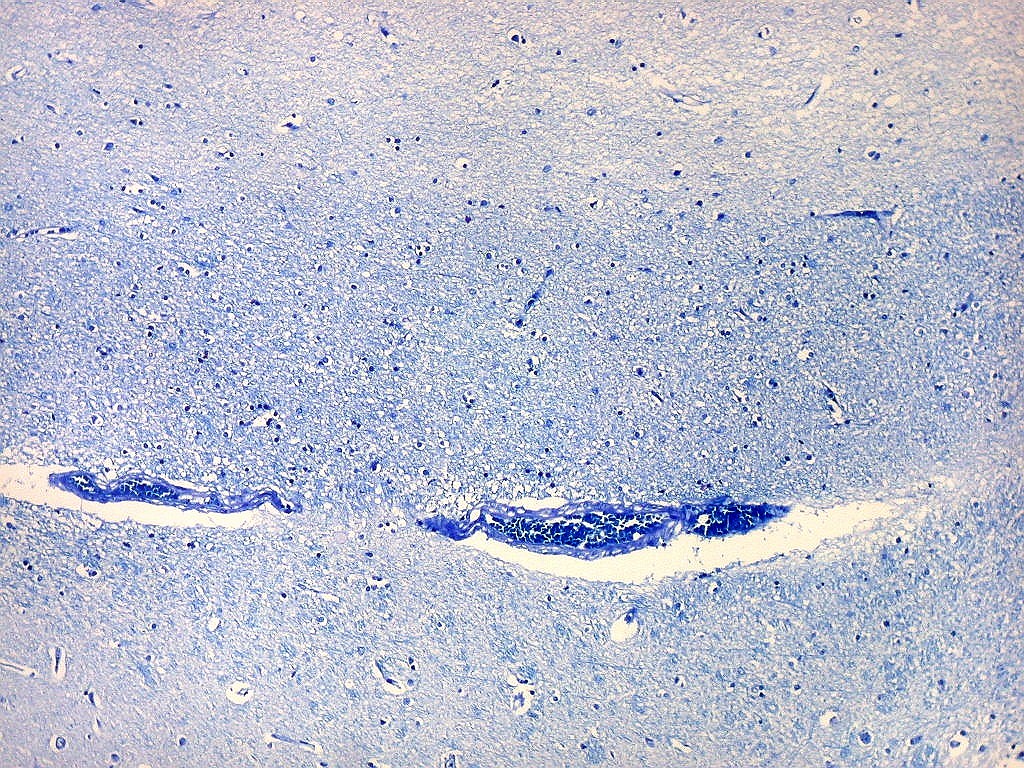 |
| Campo 7, sulco hipocampal, entre giro denteado e pró-subículo / CA1 |
 |
 |
 |
 |
| Campo 8, sulco hipocampal, extremidade lateral. |
 |
 |
 |
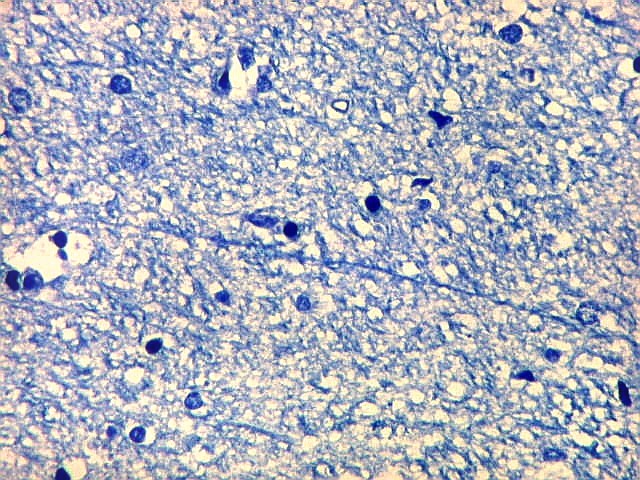
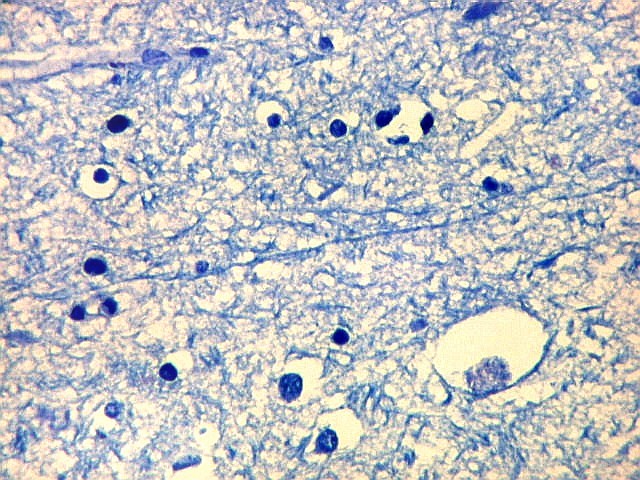
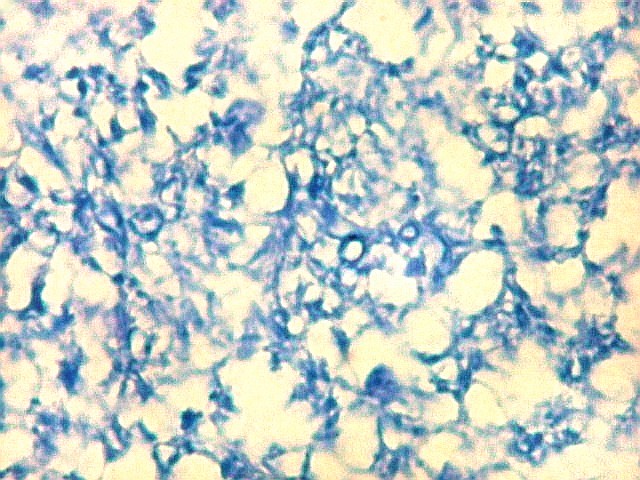
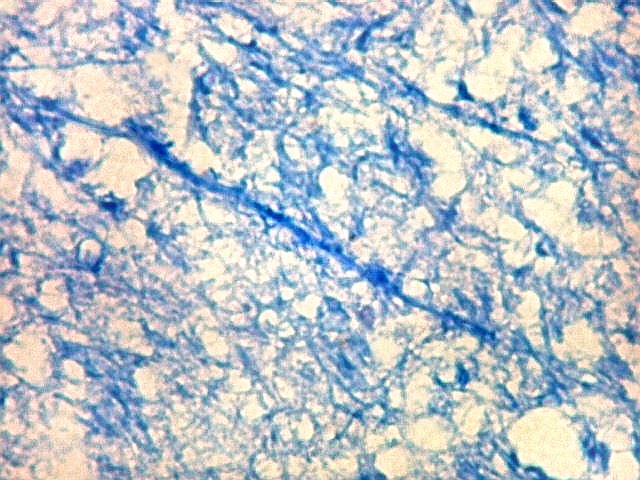
| Campo 9, CA1, alveus, ventrículo lateral. Para detalhes de CA1 e restante do córtex hipocampal, clique. |
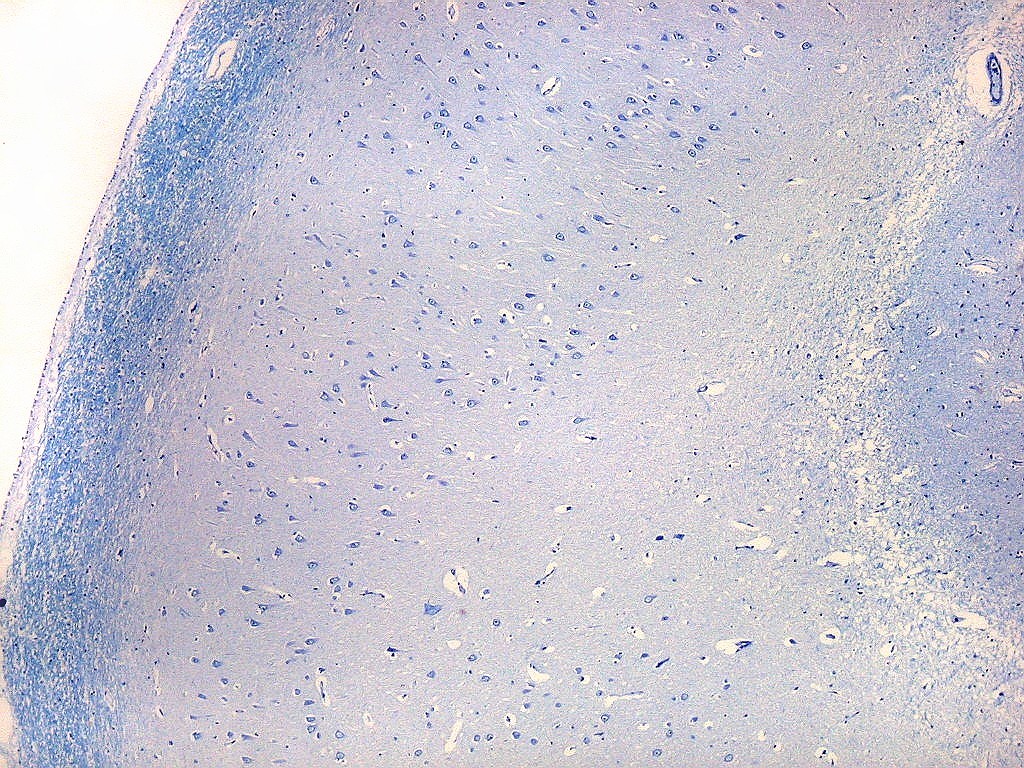 |
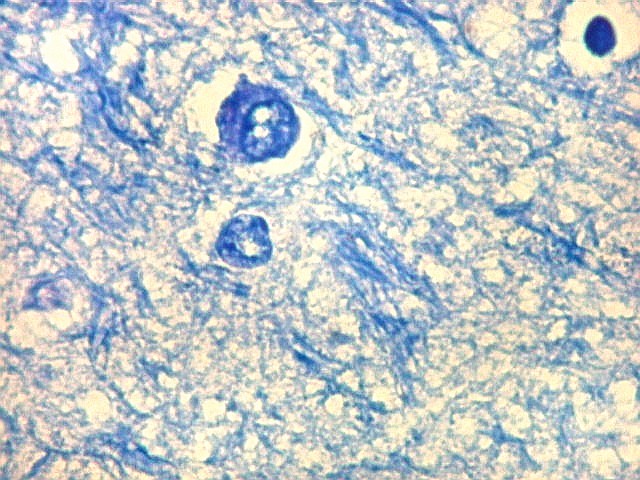
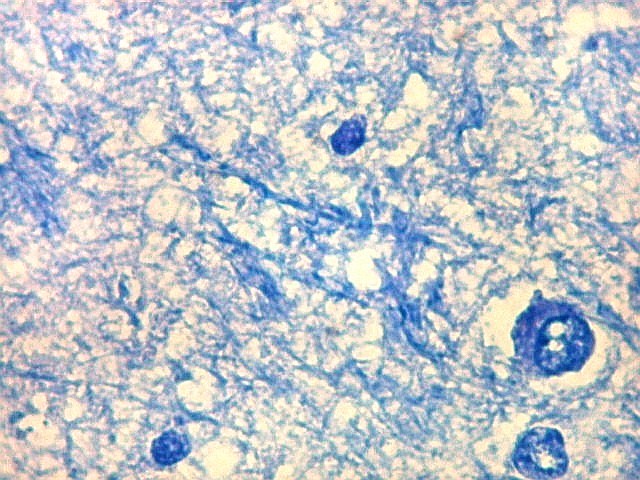
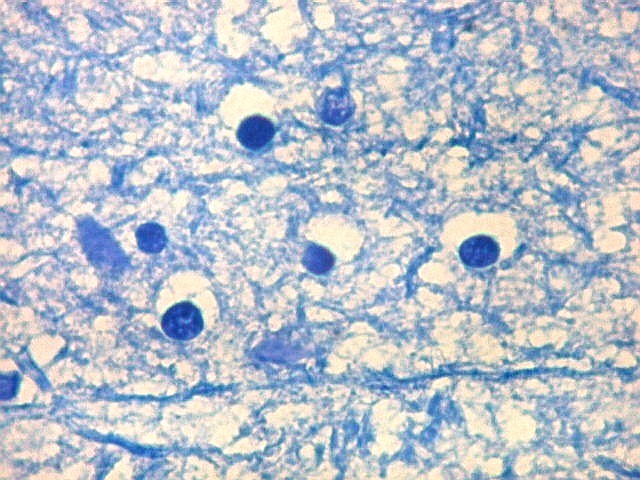
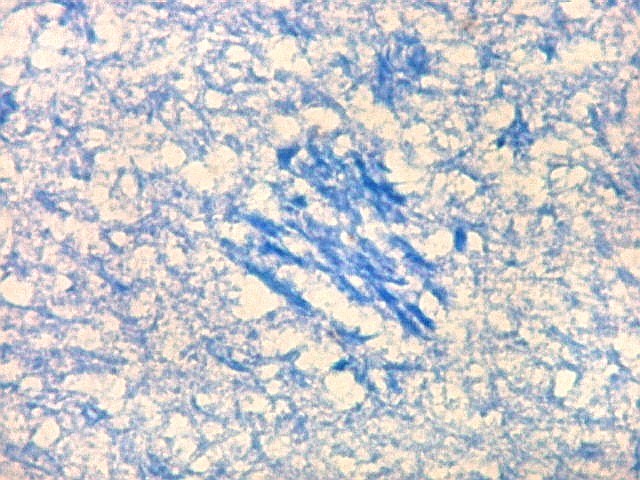
| Detalhes da substância branca ao longo do sulco hipocampal, com axônios mielínicos em corte transversal e longitudinal. | |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
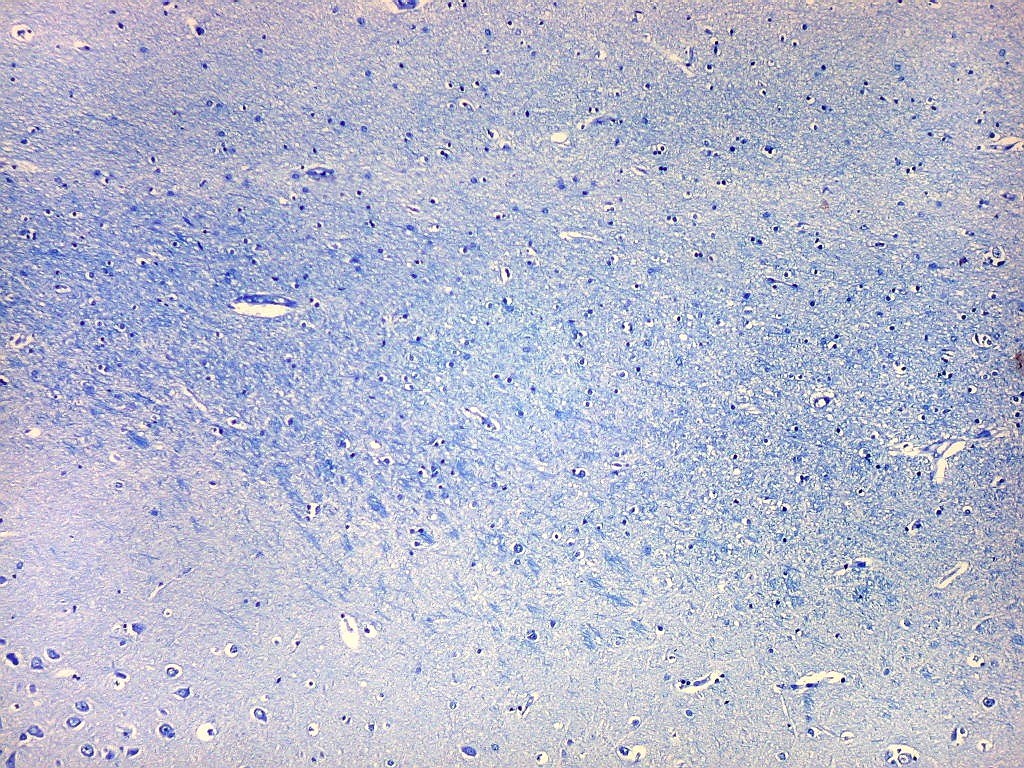
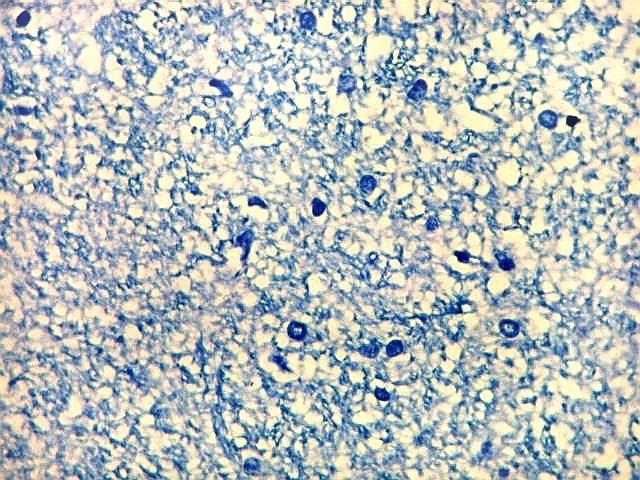
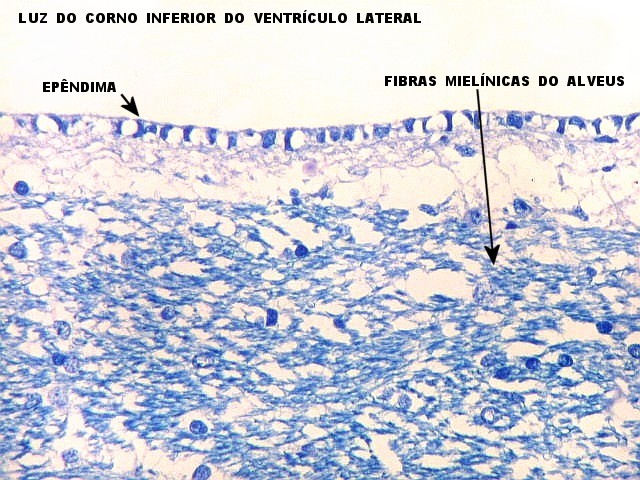
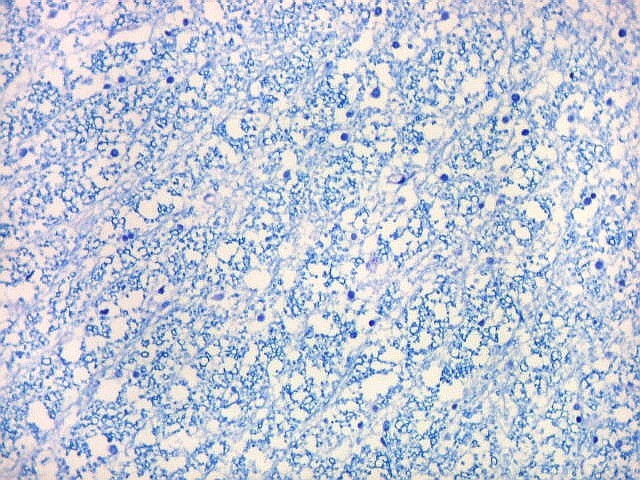
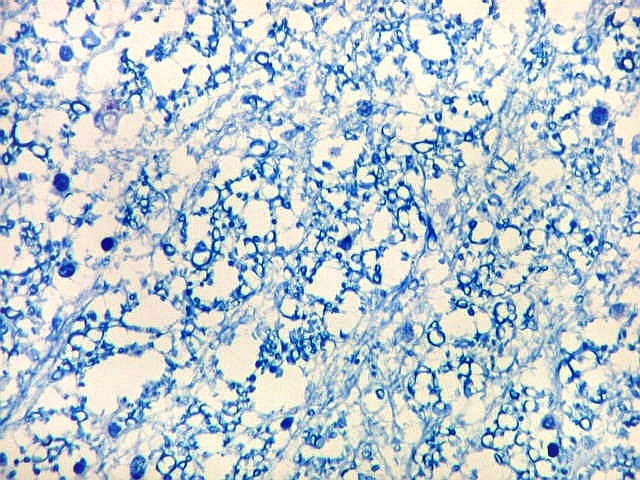
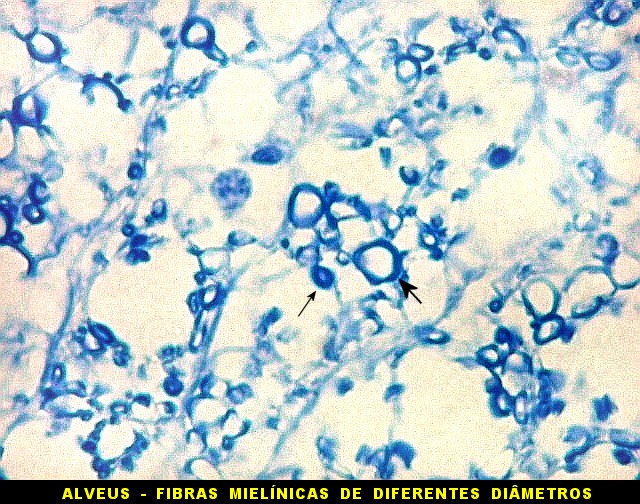
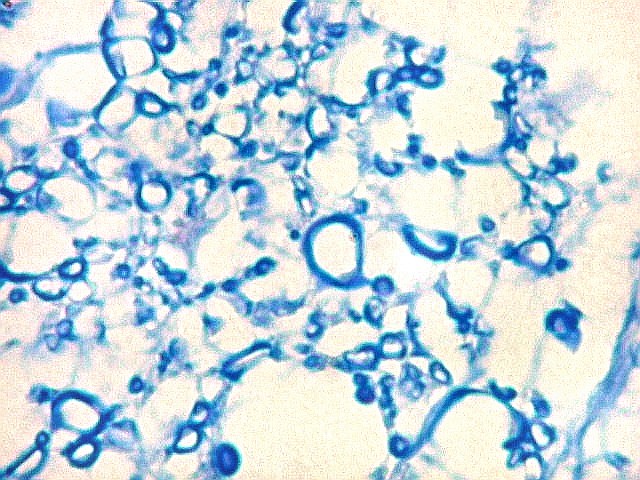
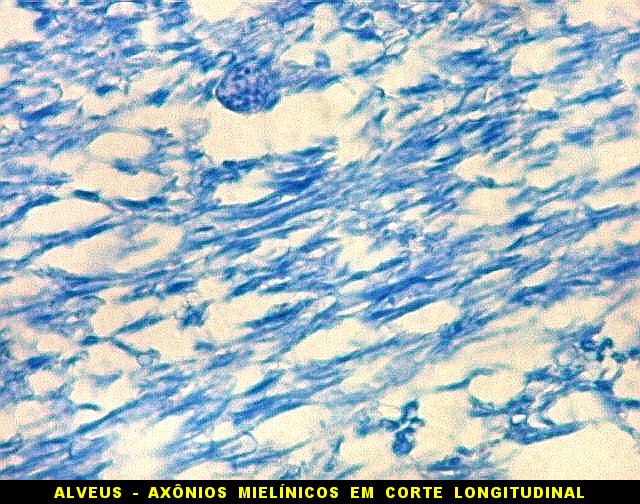
| Alveus. O alveus é a camada de substância branca que recobre a superfície ventricular do hipocampo. Como toda superfície ventricular, tem uma camada de epêndima, o neuroepitélio de células colunares que reveste todos os ventrículos cerebrais e o canal central da medula espinal. O alveus é formado por axônios mielínicos de calibre variável, na maioria pequenos, e de células da glia, como discutido abaixo na fímbria. Contém na maioria fibras eferentes do hipocampo que são reunidas na fímbria, mas há também fibras aferentes de fora do hipocampo. Para breve texto, clique. | |
|
 |
 |
 |
 |
 |
 |
 |
 |
 |
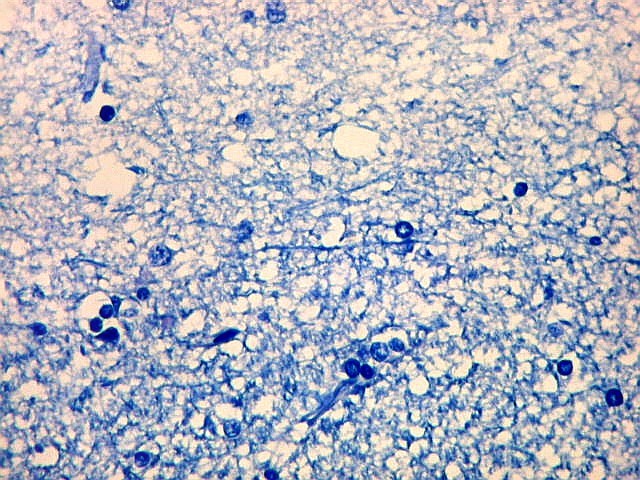
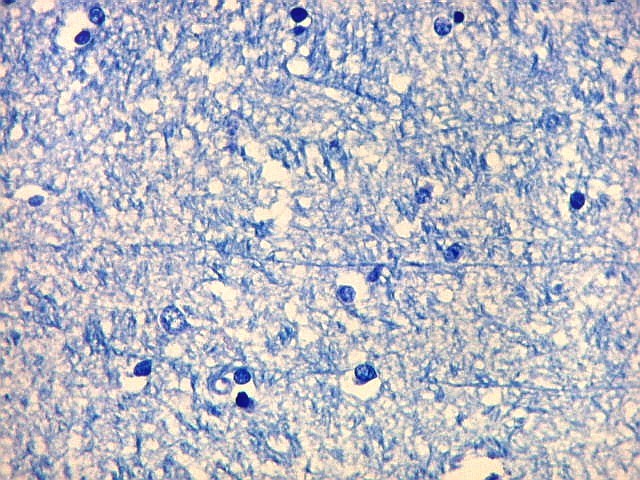
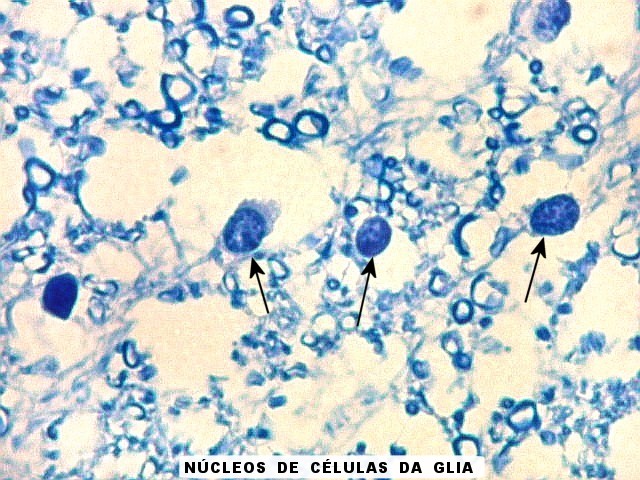
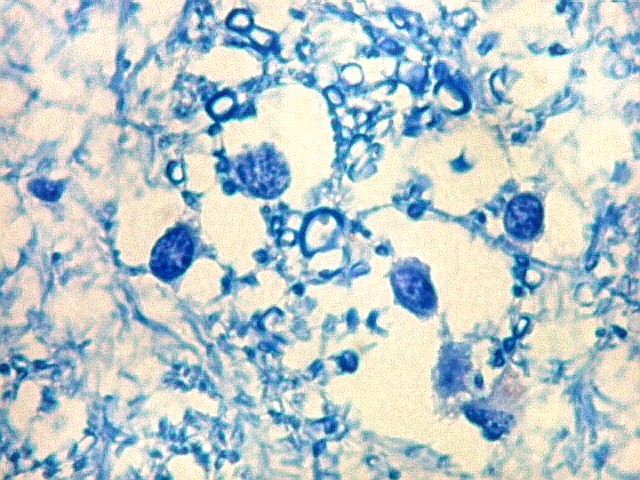
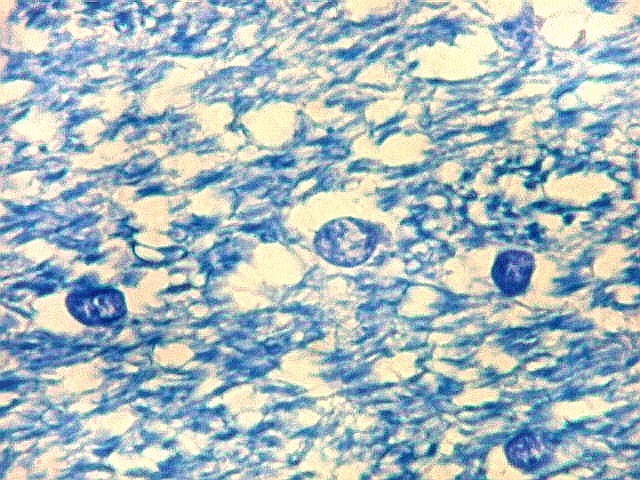
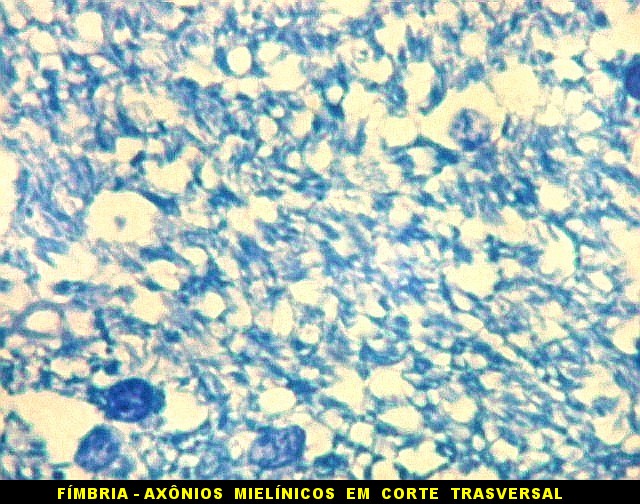
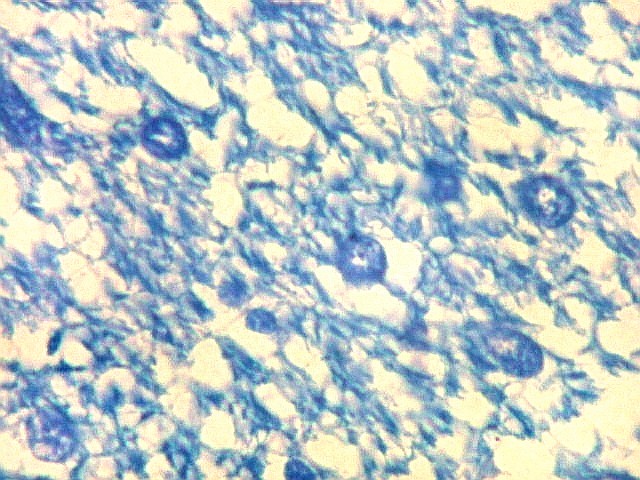
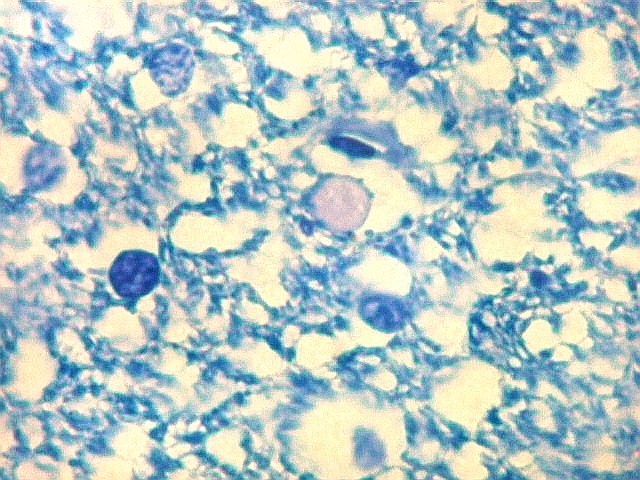
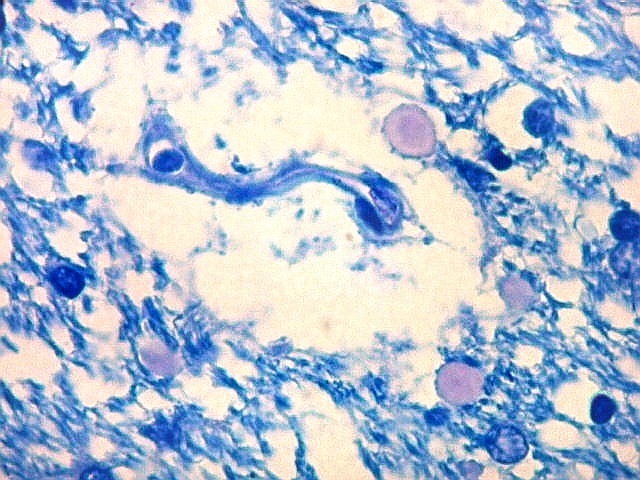
| Fímbria. Trato de substância branca que reúne axônios do alveus e se continua com o fórnix. Este dirige-se aos corpos mamilares. Para a fímbria, fórnix e corpos mamilares em peças anatômicas, clique. A fímbria é composta por axônios mielínicos e células da glia. Clique para os núcleos destas em HE, astrócitos na imunohistoquímica para GFAP, e micróglia na imunohistoquímica para CD68 e axônios em neurofilamento. Fotos em aumento forte (abaixo) mostram axônios mielínicos muito finos em corte transversal, núcleos de células da glia e alguns corpos amiláceos como pequenas esferas homogêneas e levemente róseas. Estes são também notados em HE. |
 |
 |
 |
 |
 |
 |
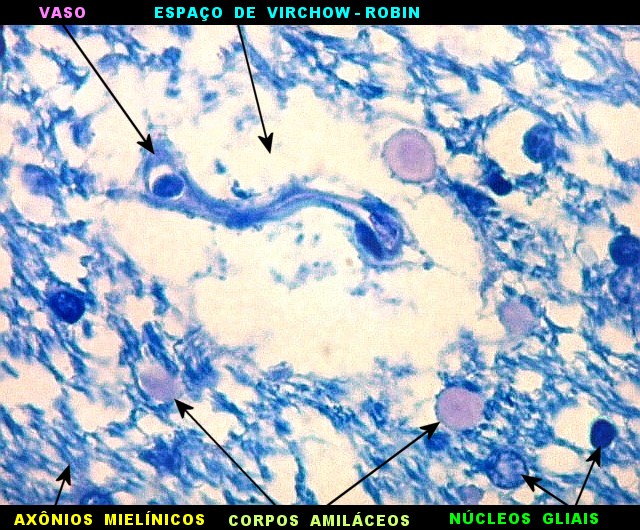 |
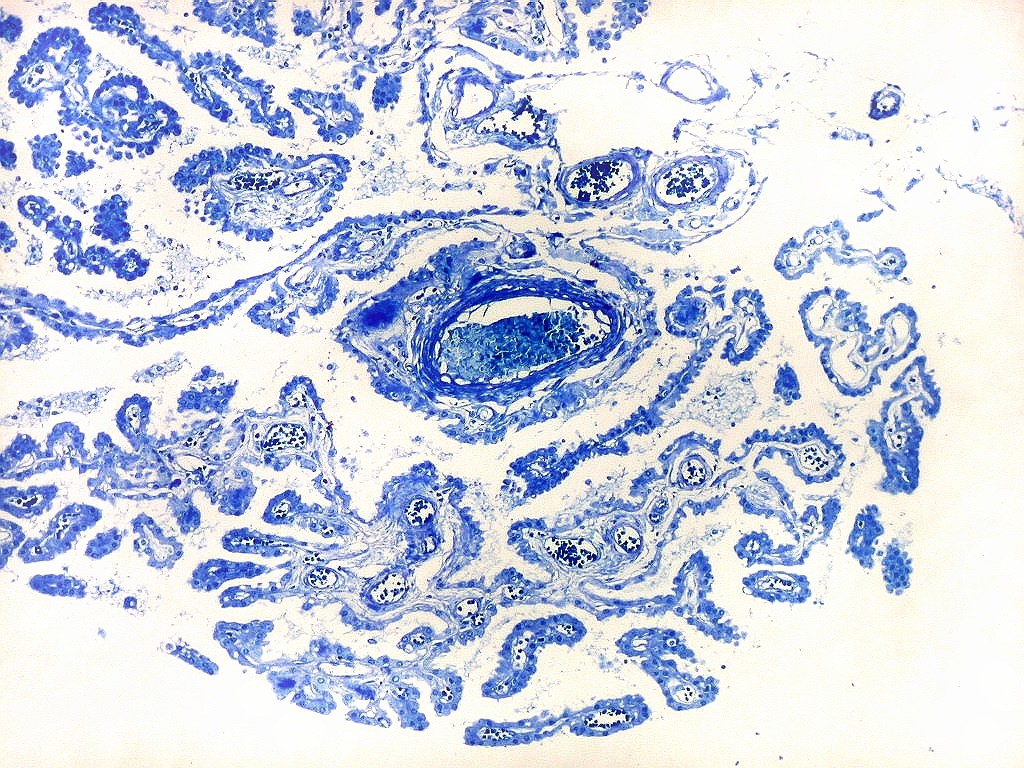
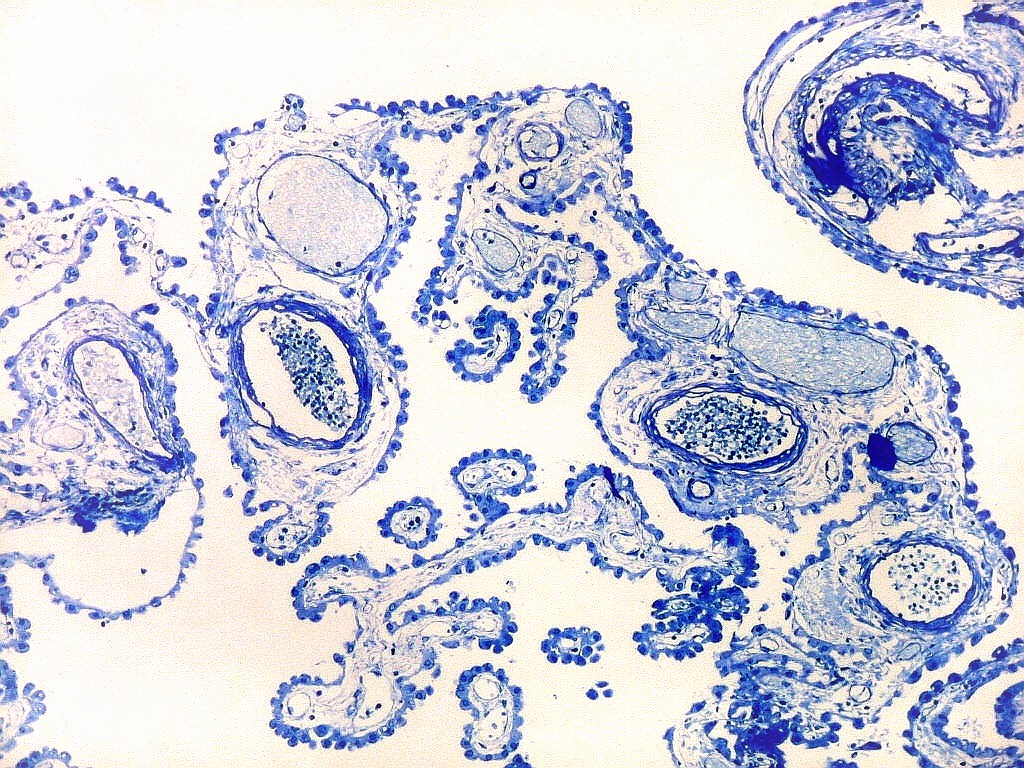
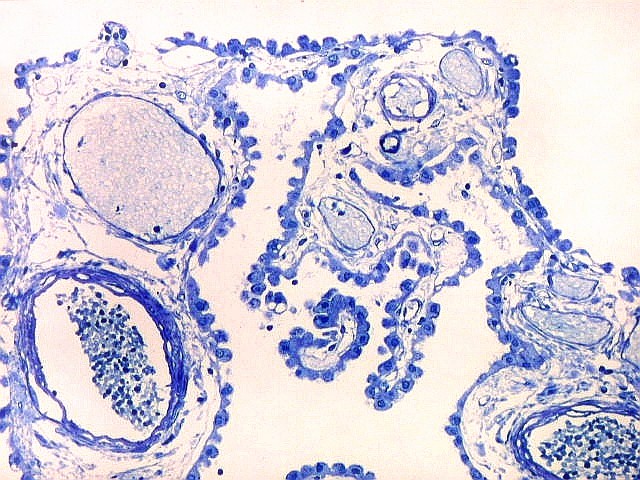
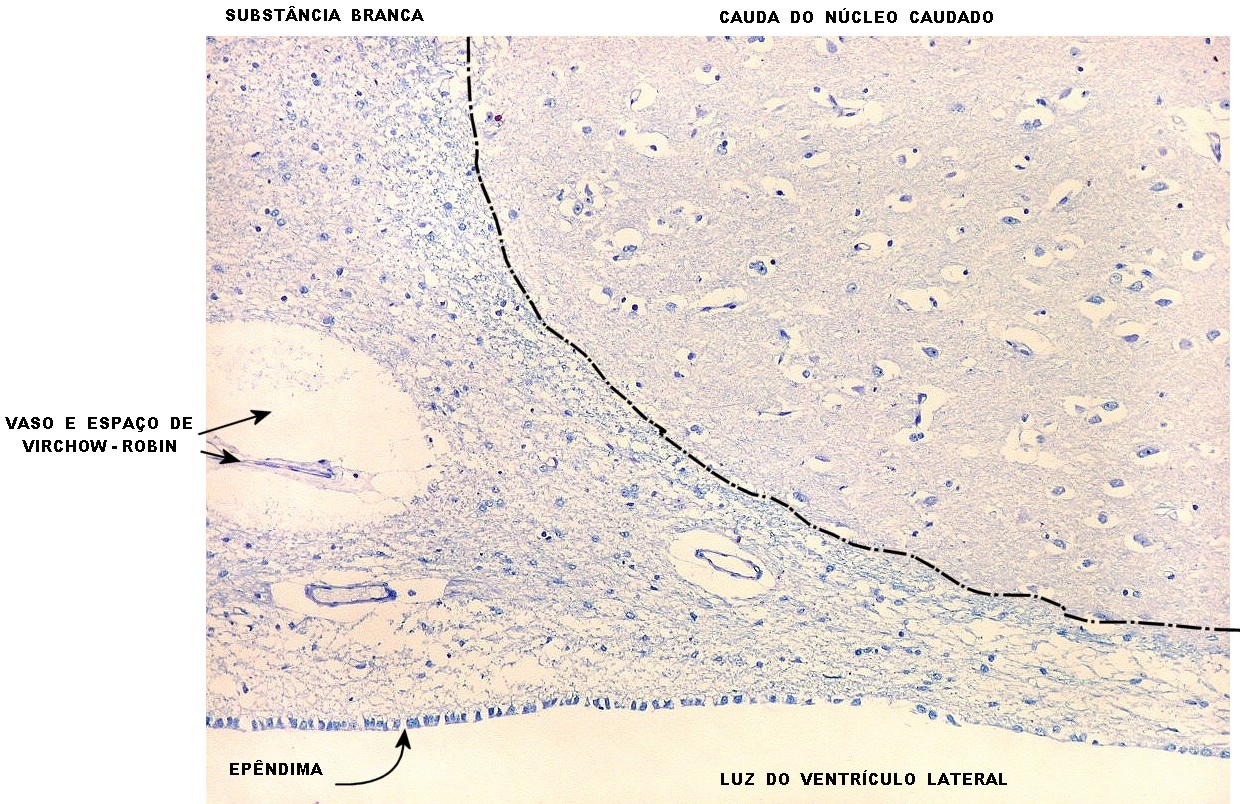
| Espaços perivasculares de Virchow-Robin são continuações em dedo de luva do espaço sub-aracnóideo e contêm líquor. São vistos ao longo de pequenos vasos próximos à leptomeninge. Podem parecer artificialmente dilatados pela perda de pressão nos vasos após a retirada do espécime cirúrgico, ou após a morte, em material de autópsia. A retração do tecido na inclusão em parafina contribui para o alargamento dos espaços de Virchow-Robin. São importantes como via de penetração de exsudato e microorganismos nas meningites bacterianas ou por fungos, como na criptococose. | |
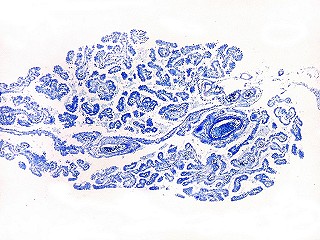
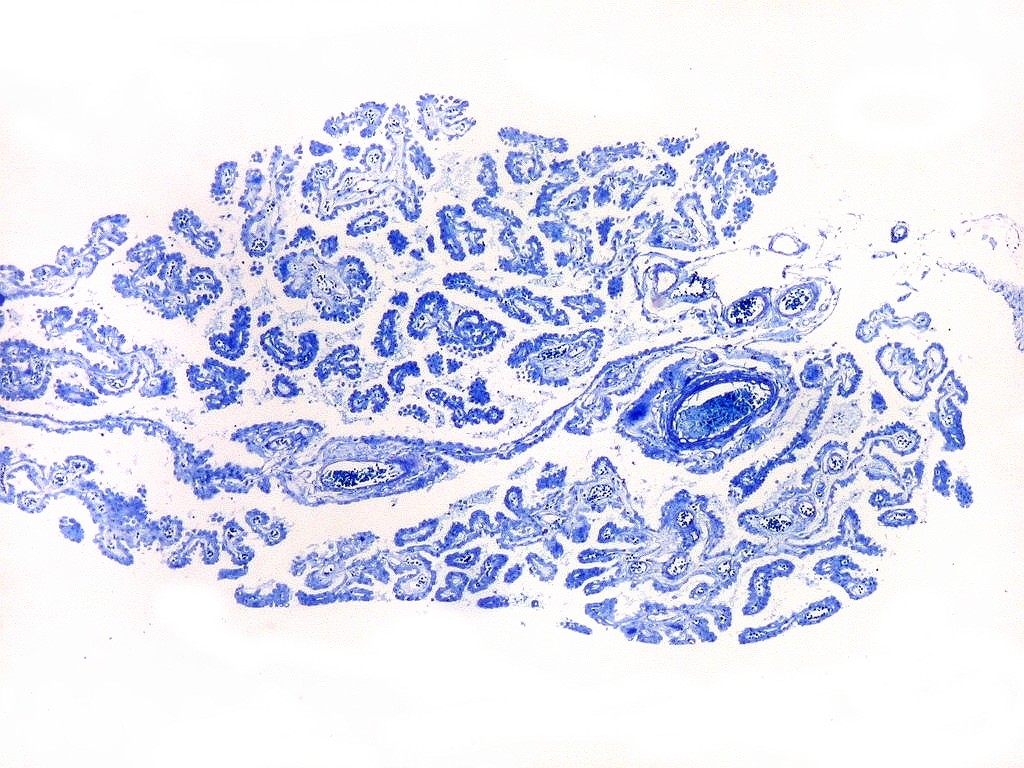
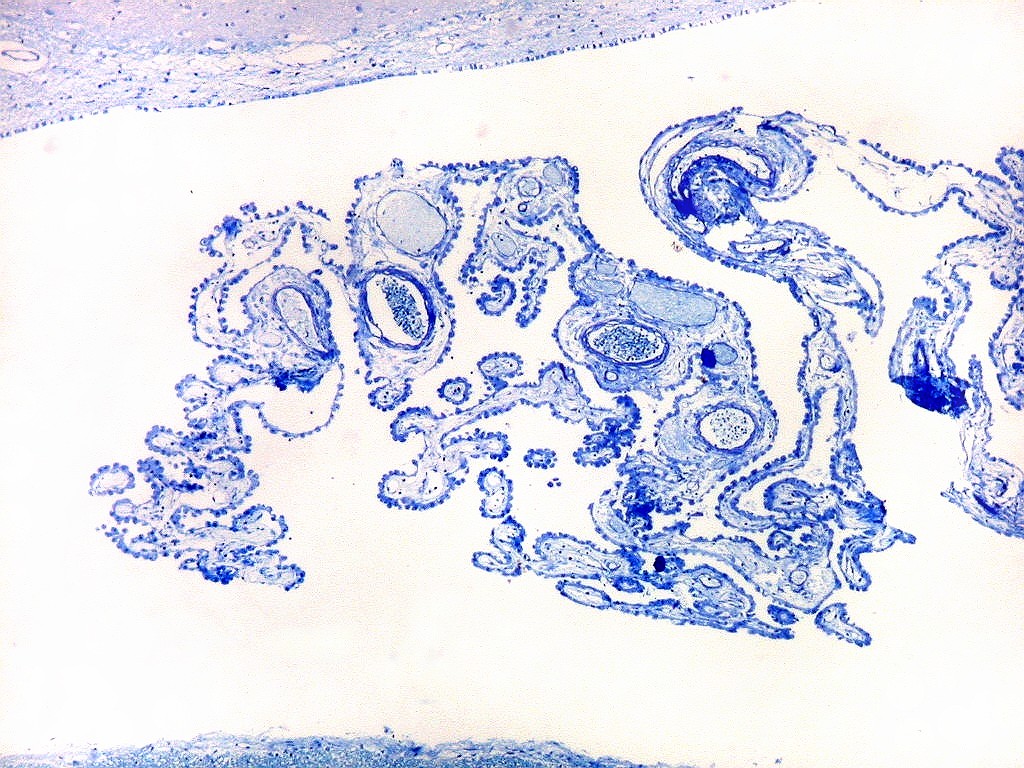
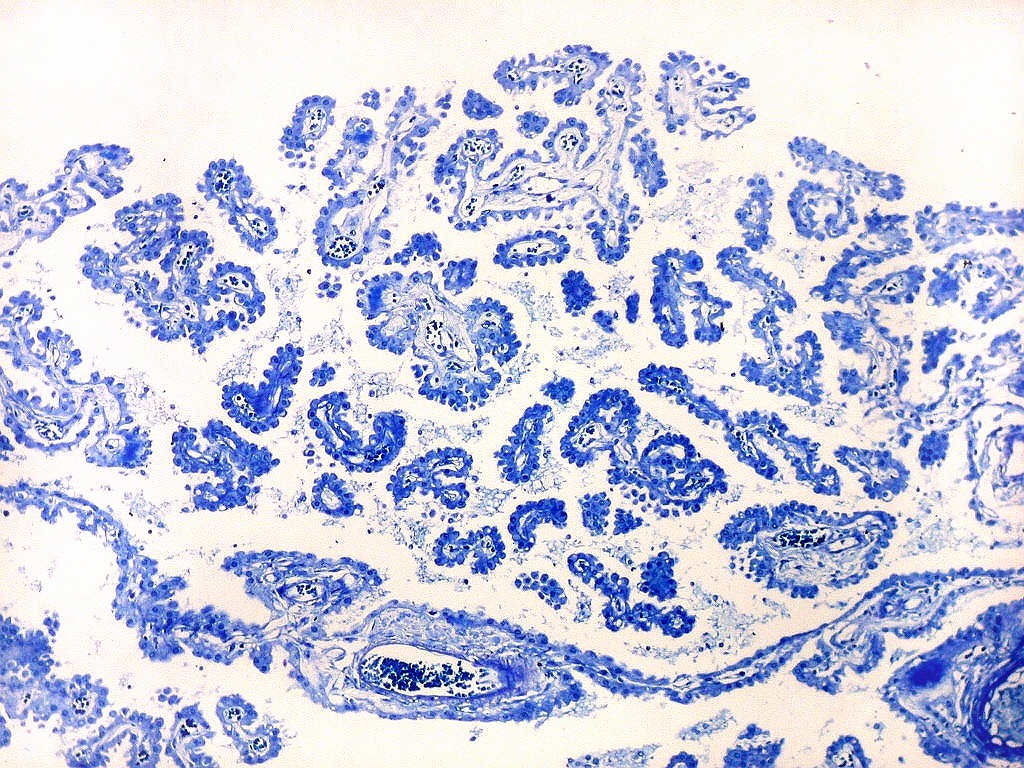
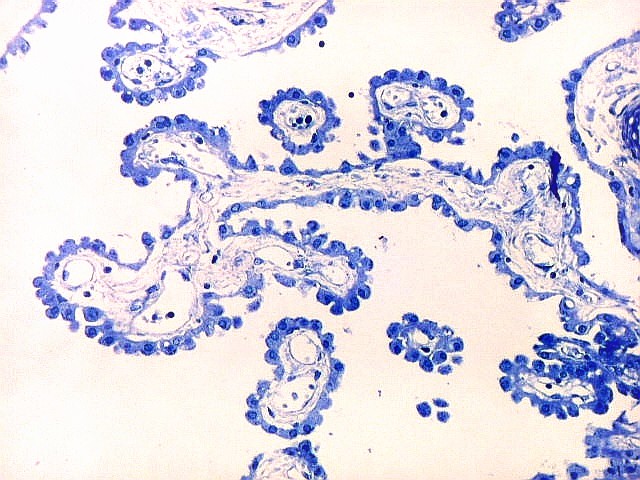
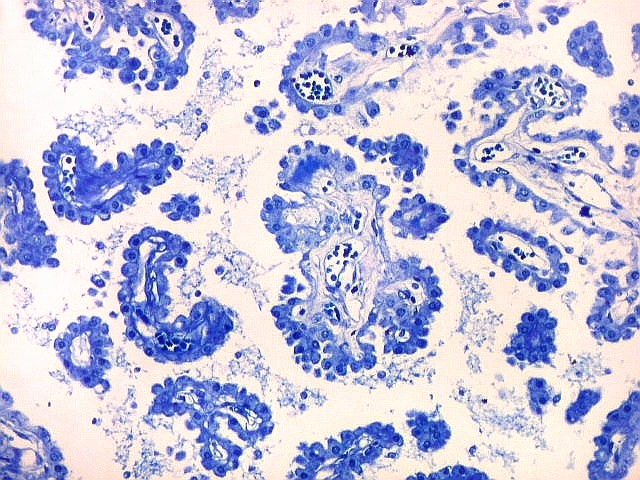
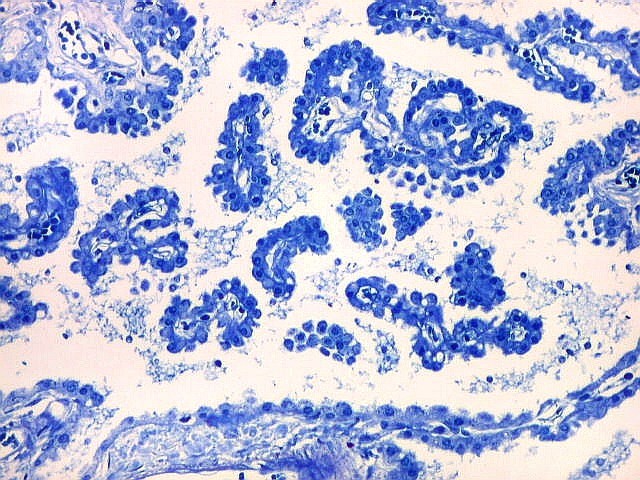
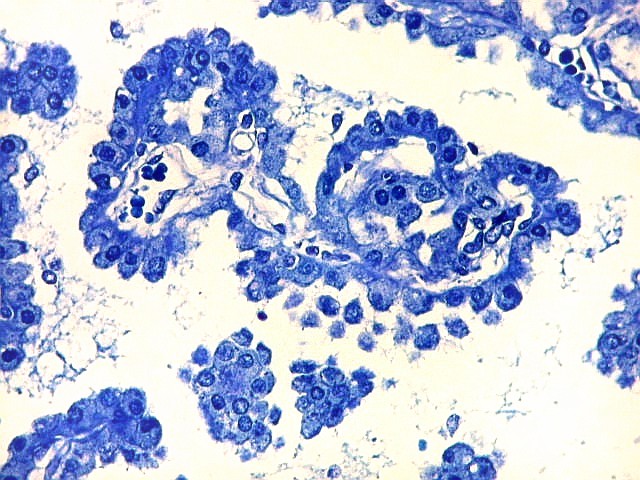
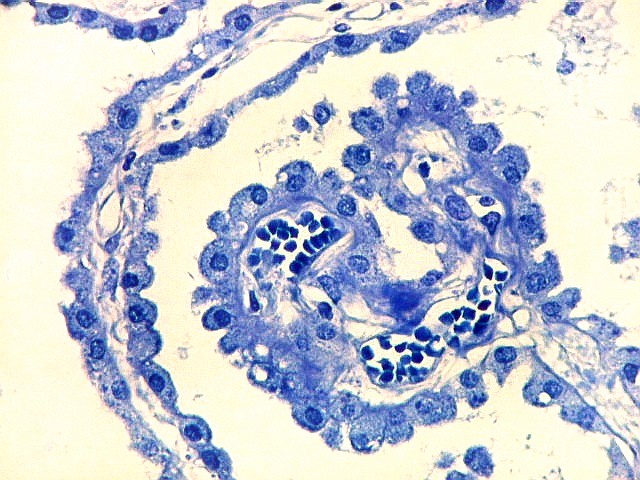
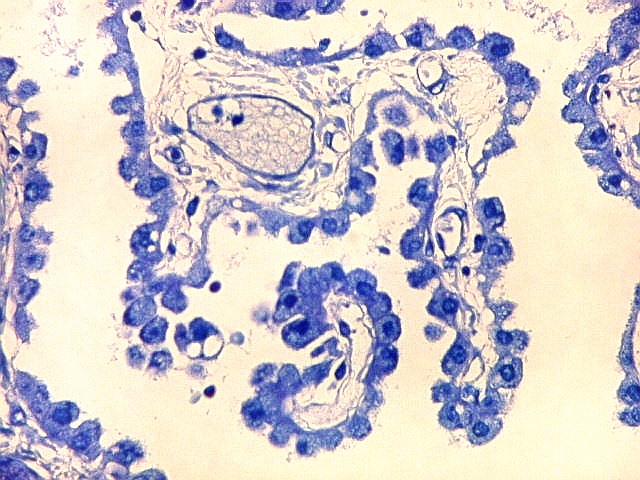
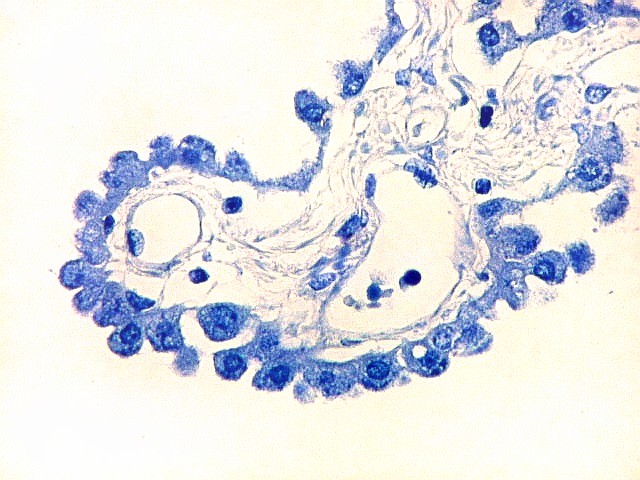
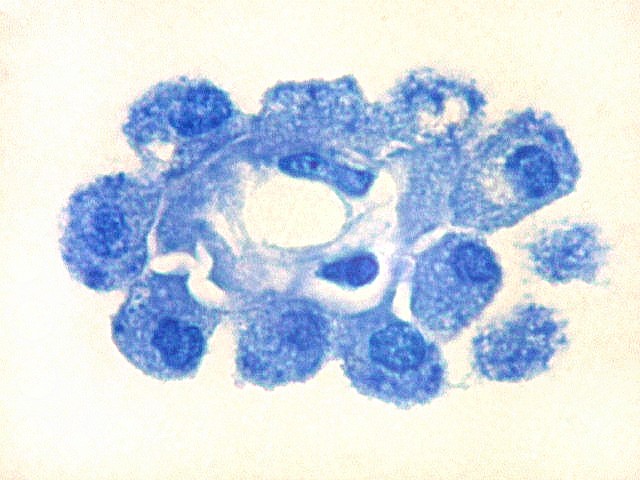
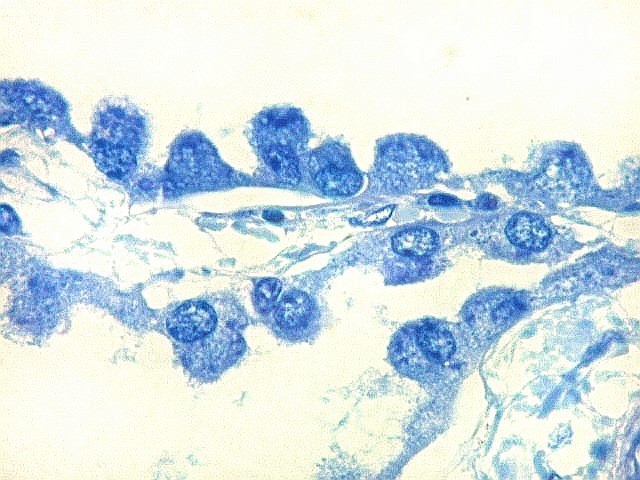
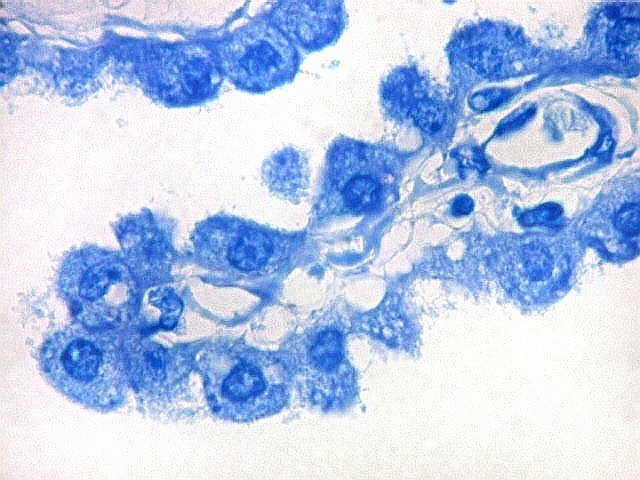
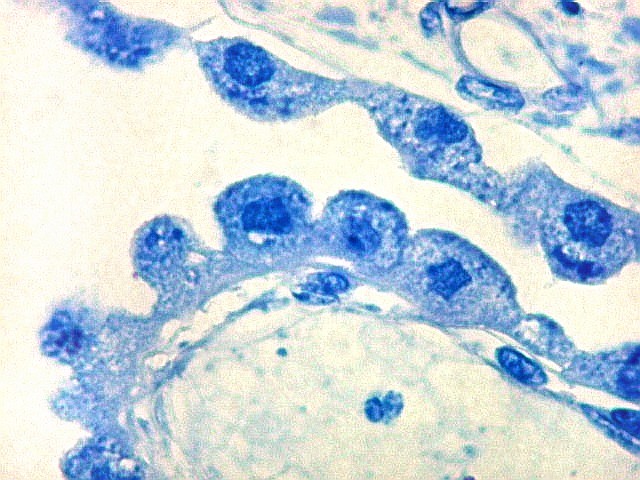
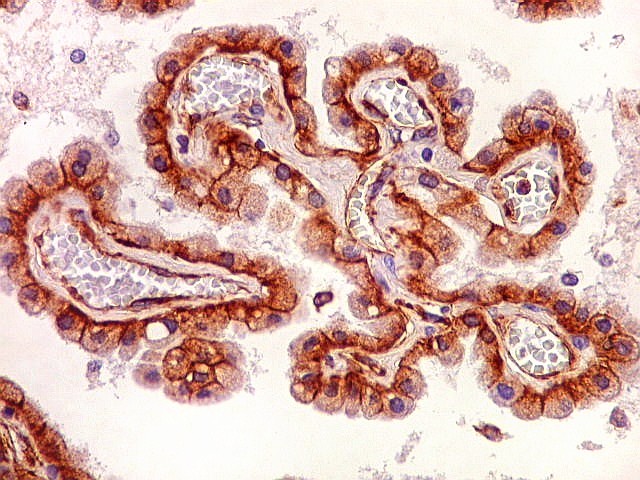
| Plexo coróide. Esta glândula produtora de líquido céfalo-raquidiano (líquor) está discutida em detalhe na página de HE. Aqui a documentamos pelo aspecto estético, com suas papilas revestidas por monocamada de células cúbicas de superfície cupuliforme 'em lápides tumulares' demonstrada em vários tons de azul. Obviamente não há axônios mielínicos no plexo coróide, e o corante que tinge a estrutura é o cresil violeta. Para o plexo coróide em outras técnicas, clique. |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
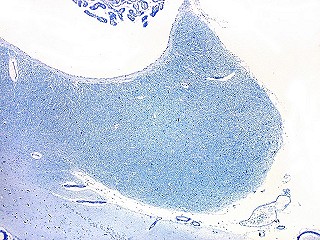
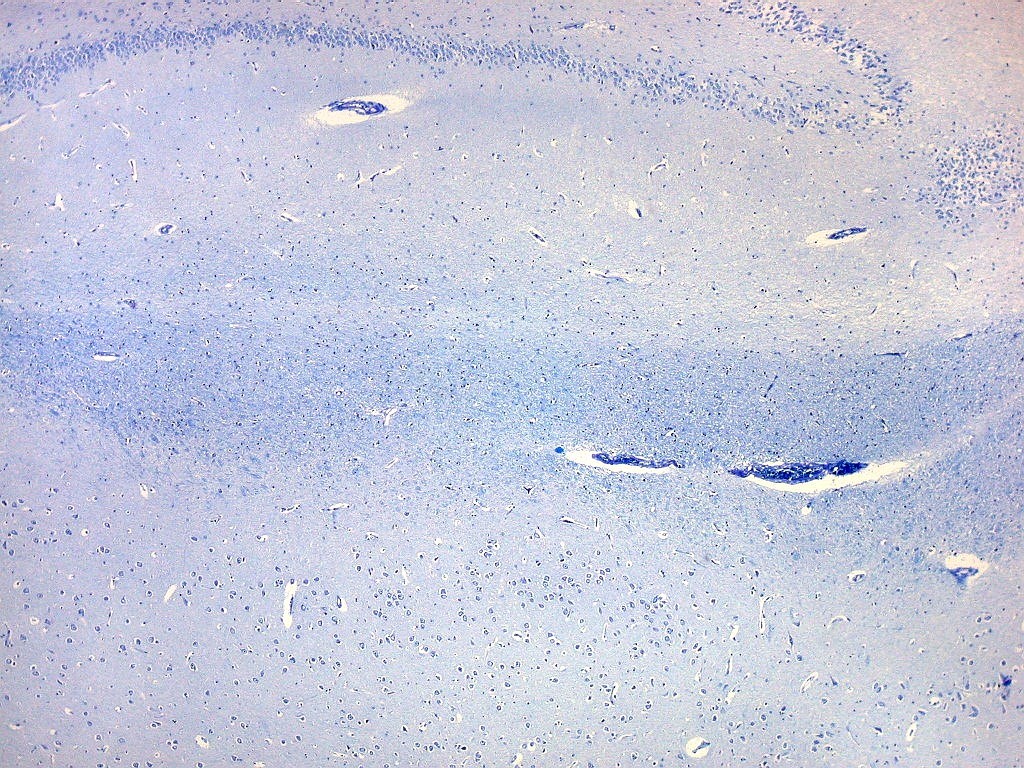
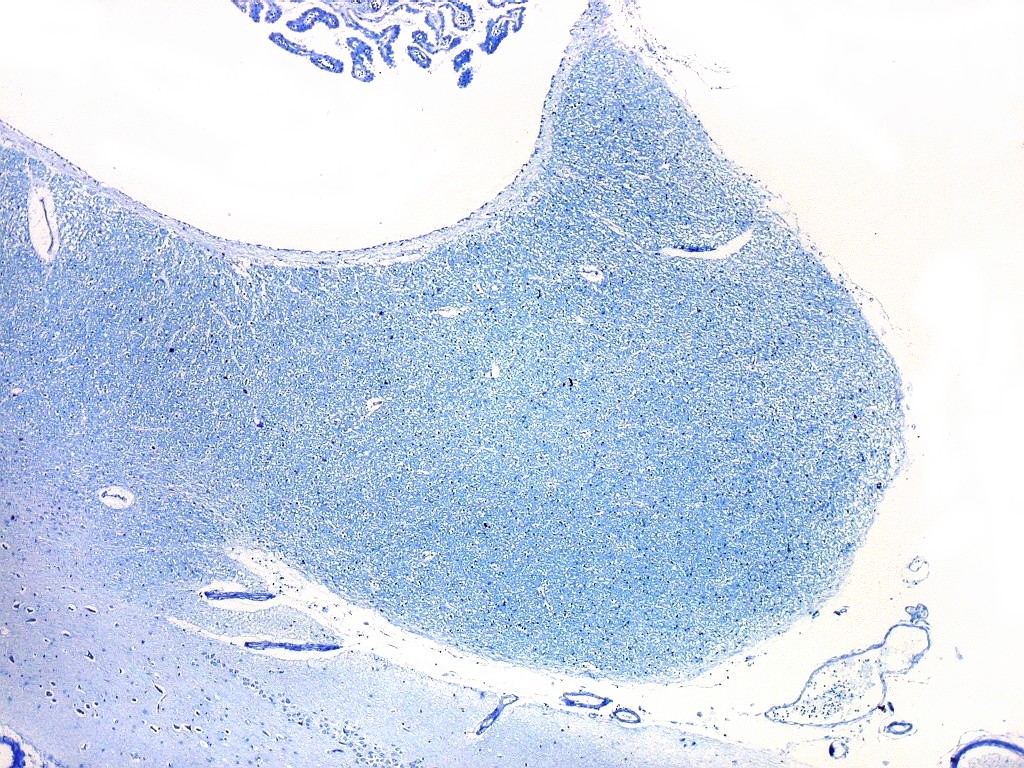
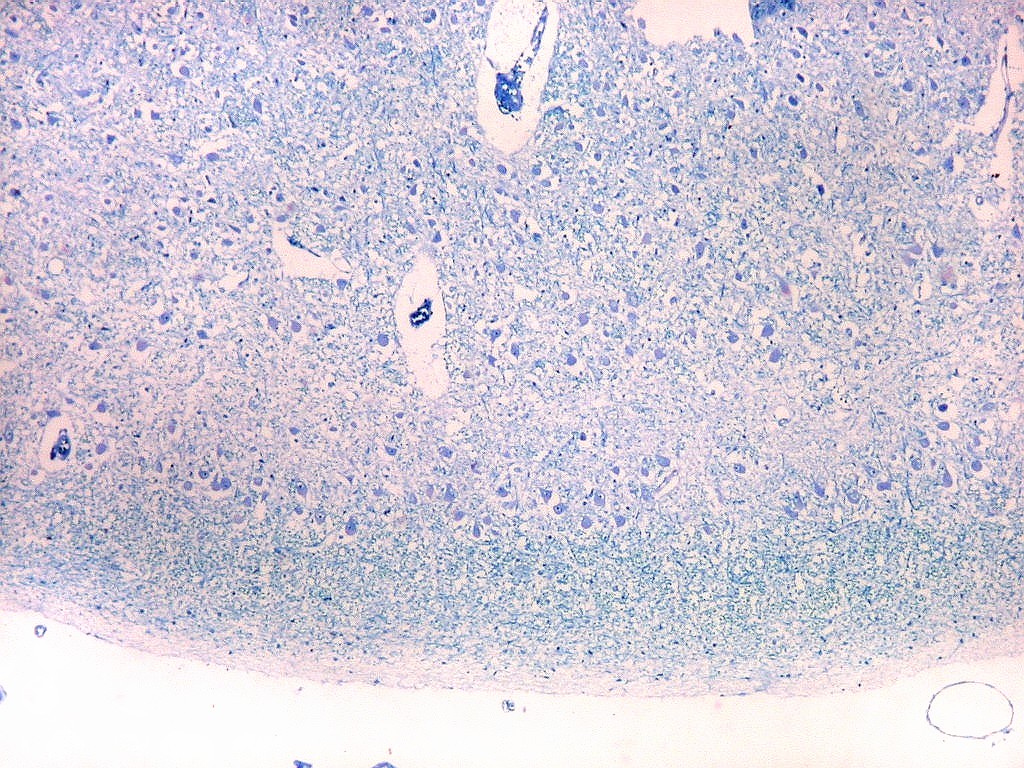
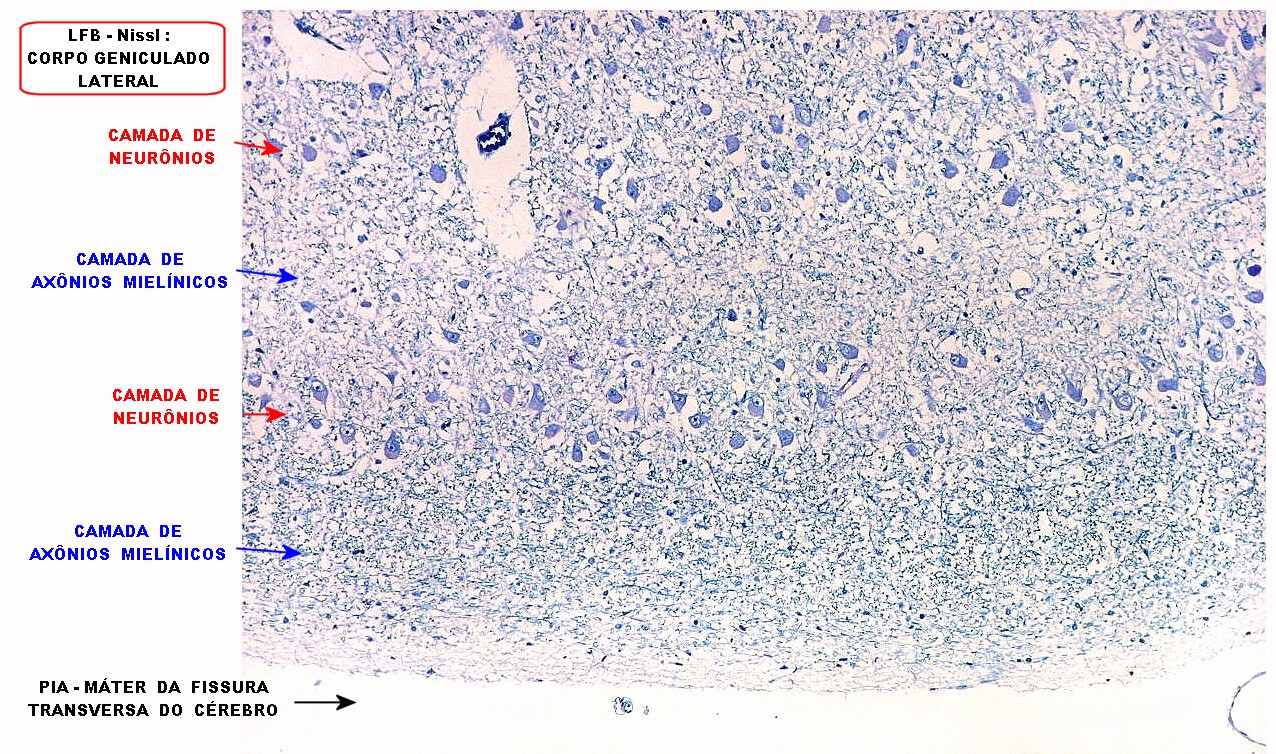
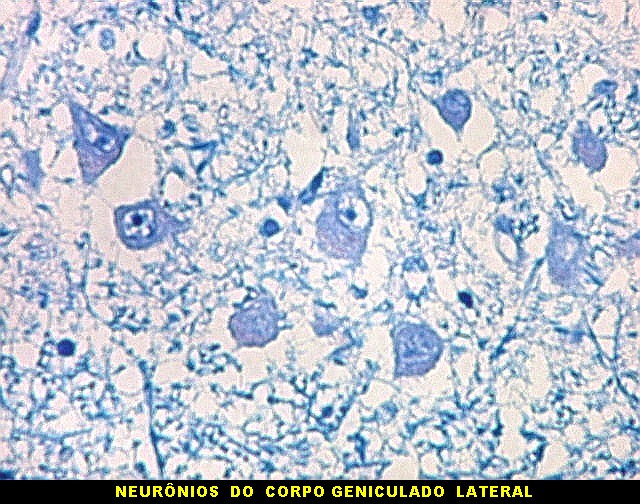
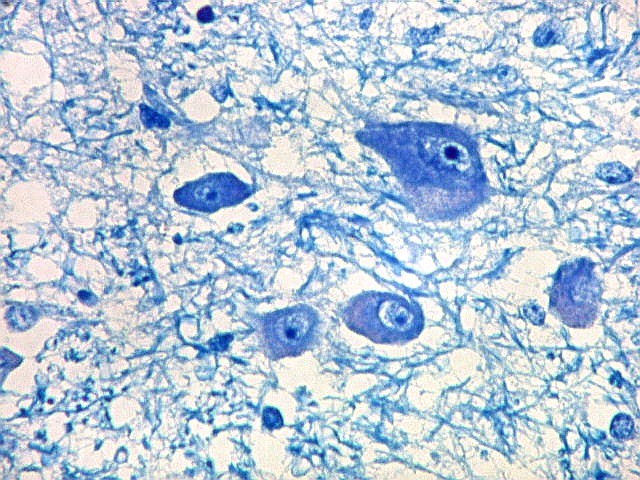
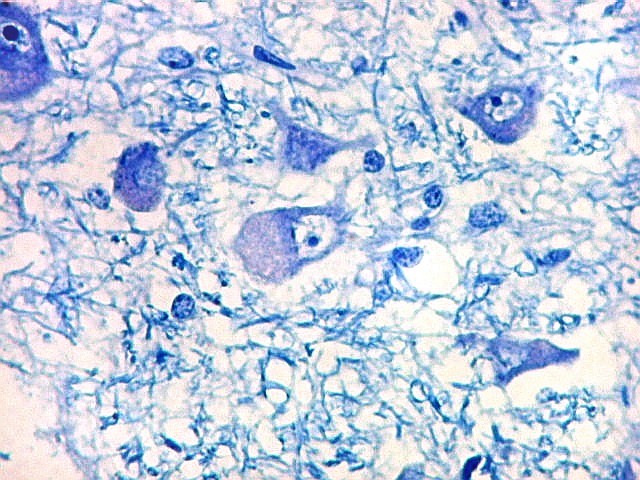
| Corpo geniculado lateral. Este núcleo da via óptica, situado medialmente à formação hipocampal na região basal do diencéfalo, alterna camadas ricas em neurônios e outras pobres, onde se destacam os axônios mielínicos demonstrados pela coloração luxol fast blue. Os neurônios são de contorno globoso e acumulam lipofuscina com a idade, assim o corpo geniculado lateral tem macroscopicamente padrão estratificado, alternando faixas brancas e pardacentas. A estrutura em camadas é também visível em cortes de celoidina. Para exemplos, clique. Para o corpo geniculado lateral em outras técnicas, clique. |
 |
 |
 |
 |
 |
|
 |
 |
 |
 |
 |
 |
 |
 |
 |
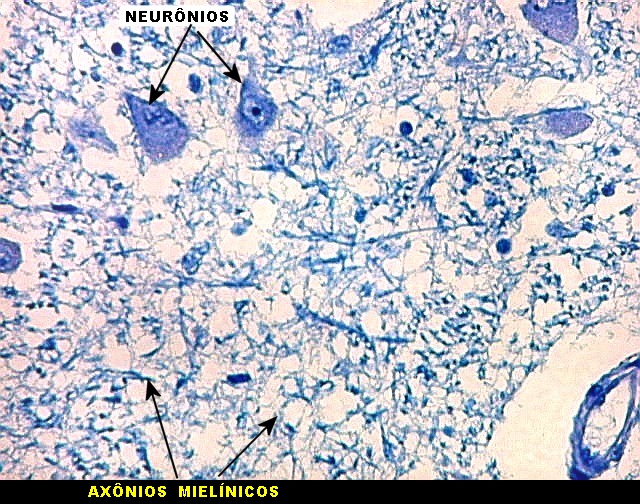
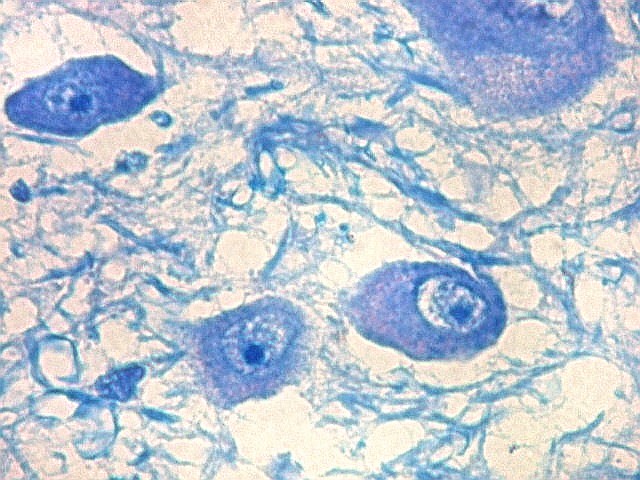
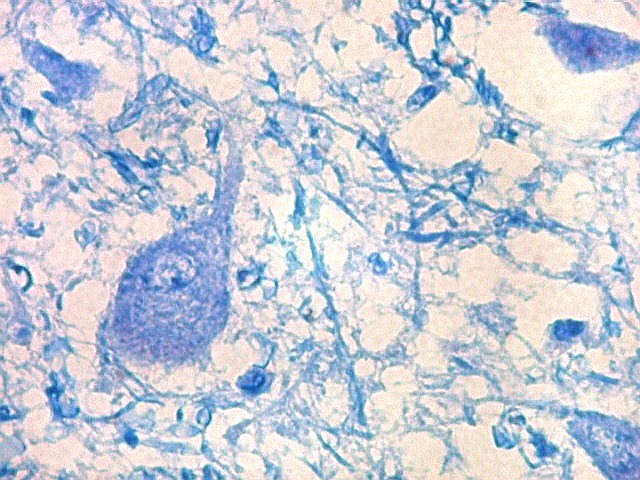
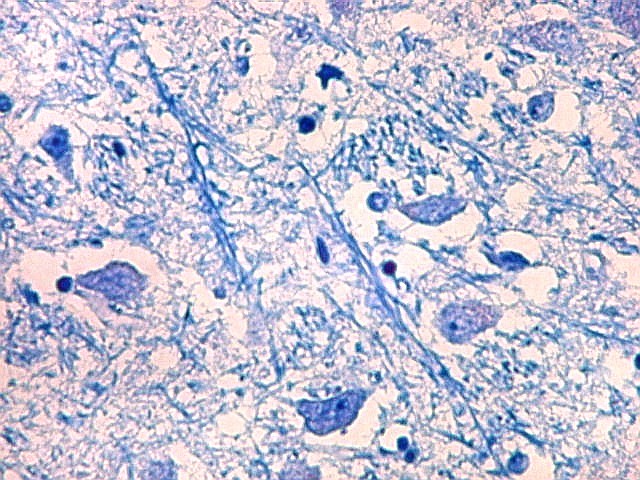
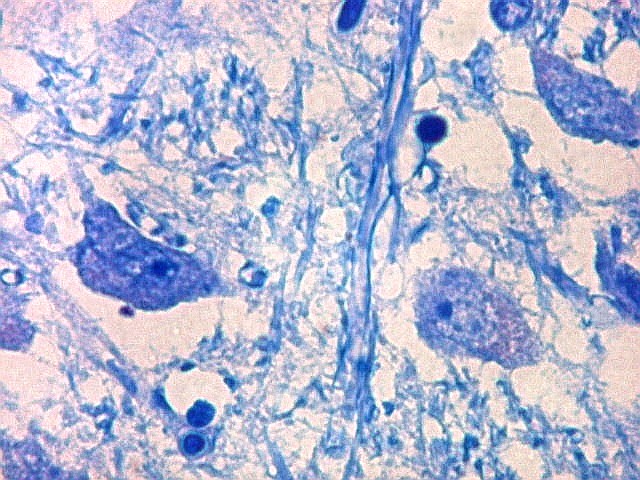
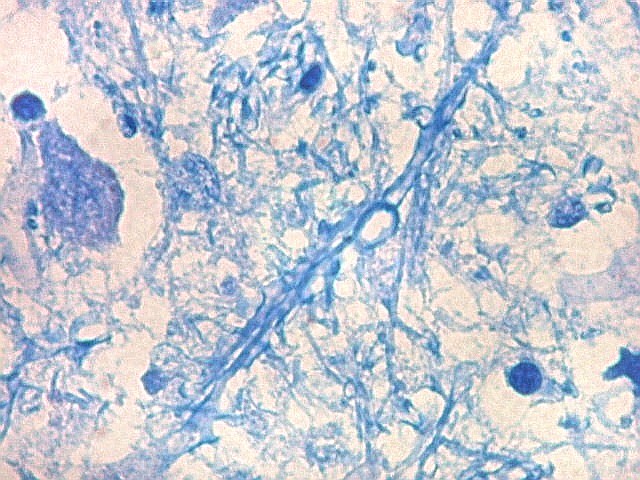
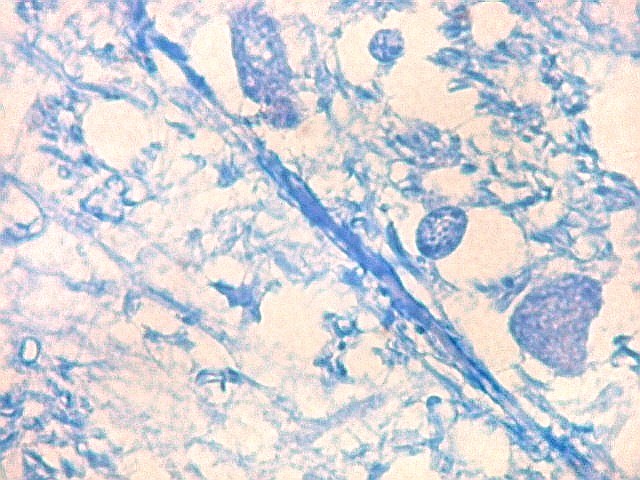
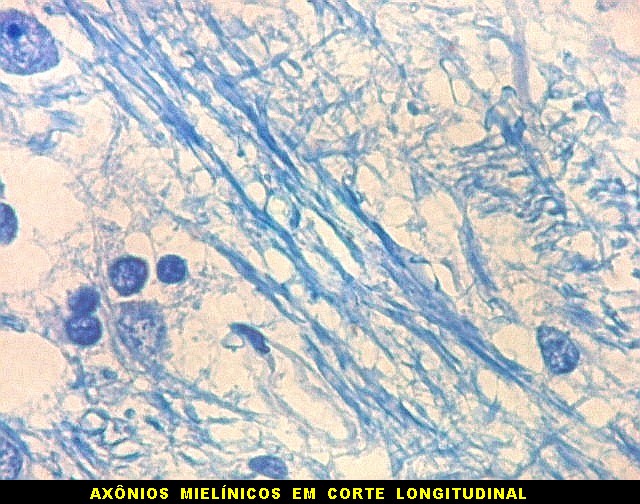
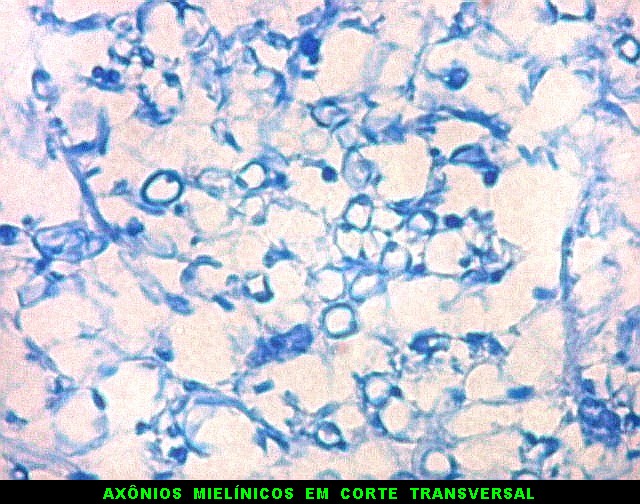
| Axônios mielínicos. A baínha de mielina é corada em azul turquesa pelo LFB e destaca-se como pequenos tubos, isoladamente ou em feixes. Em corte transversal, observa-se a luz, que aparece vazia, pois os axônios não se coram nem pelo LFB nem pelo cresil violeta (que cora os núcleos e corpos celulares dos neurônios). | |
 |
 |
 |
 |
 |
 |
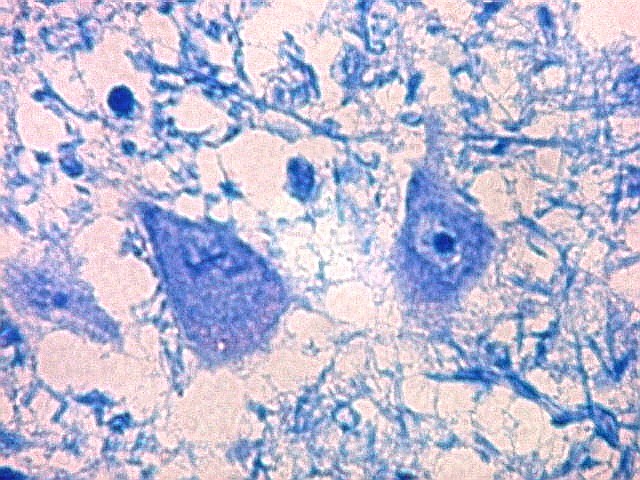
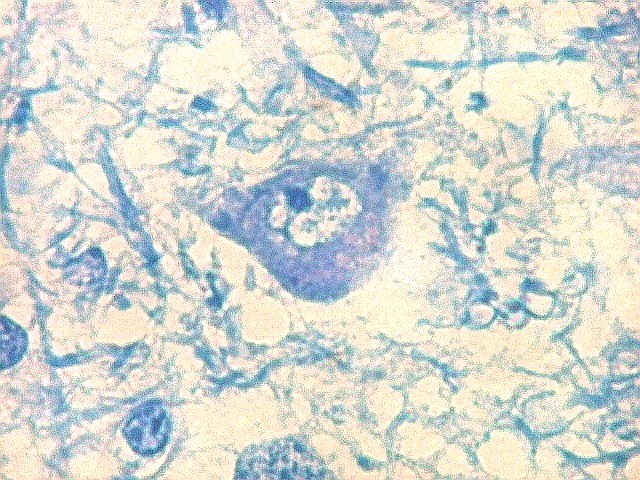
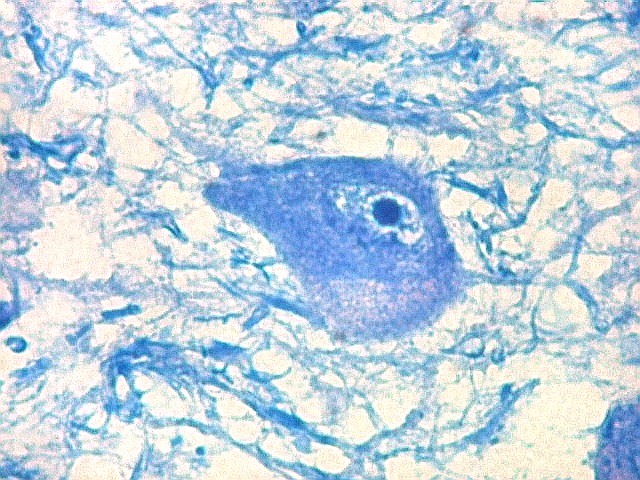
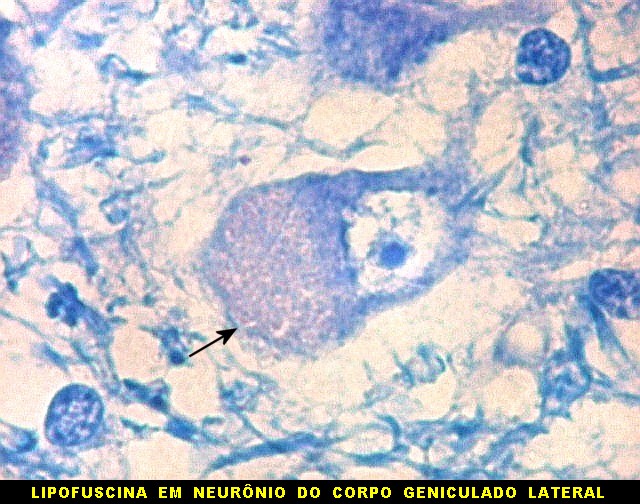
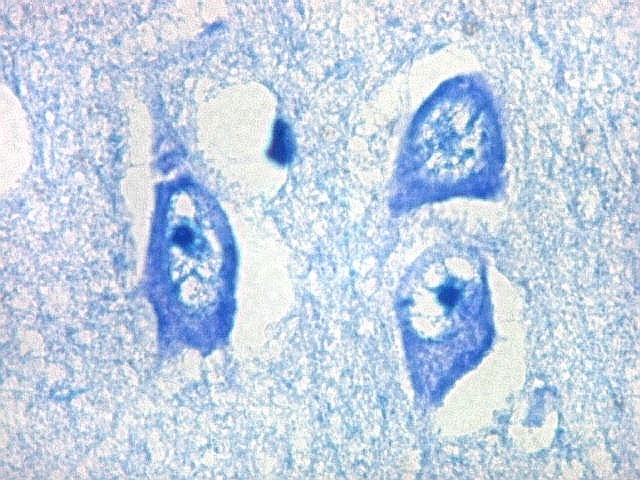
| Lipofuscina. Este pigmento de desgaste acumula-se com o avançar da idade em um setor do citoplasma destes neurônios do corpo geniculado lateral, com tonalidade levemente pardo-amarelada e aspecto granuloso. Ver em outros locais e técnicas (1)(2)(3)(4). | |
 |
 |
 |
 |
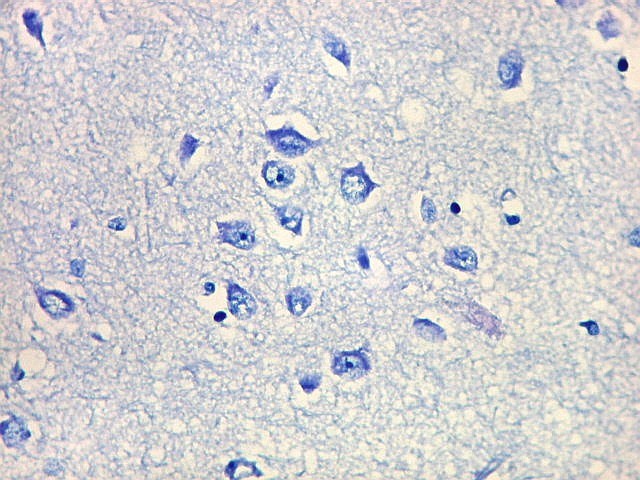
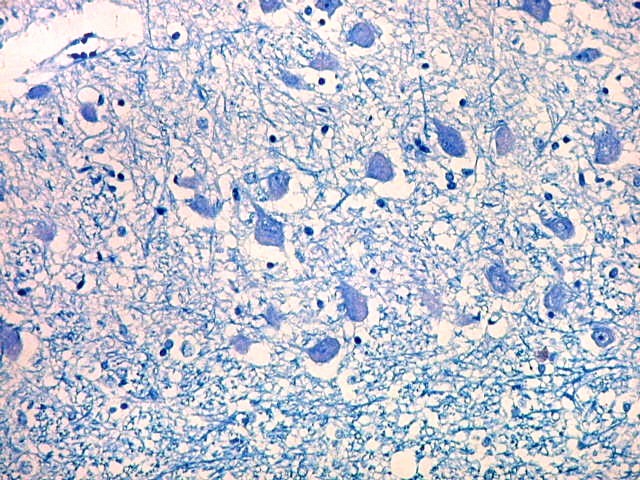
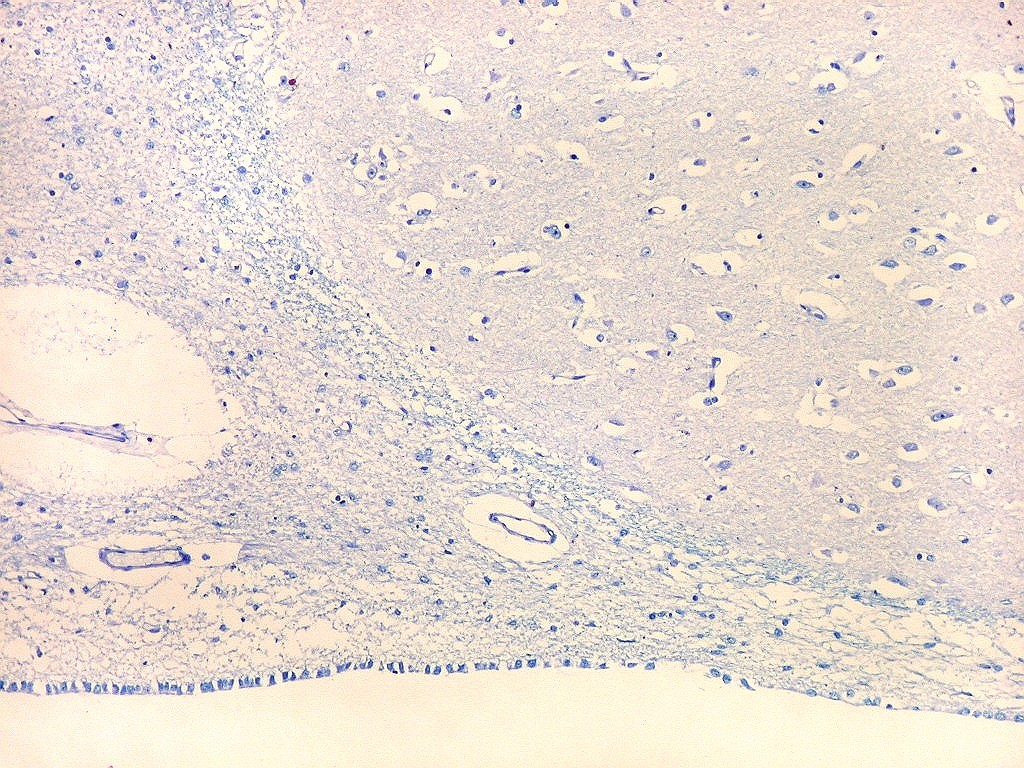
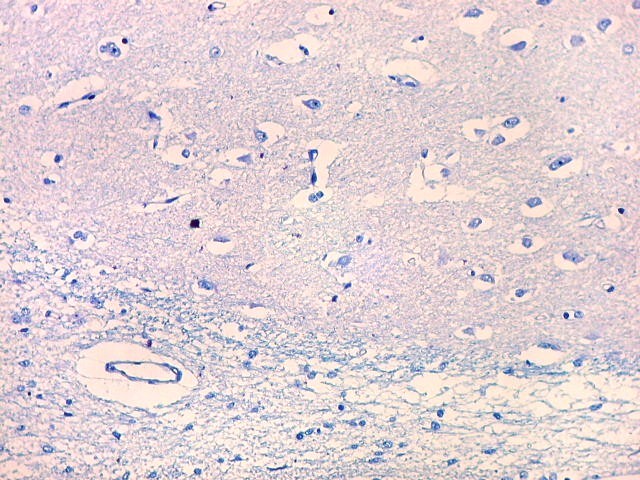
| Cauda do núcleo caudado. Esta extensão da massa principal do núcleo caudado, um dos núcleos da base do telencéfalo, situa-se no teto do corno interior do ventrículo lateral, no lobo temporal, sendo visível em cortes coronais que incluem a porção média da formação hipocampal. Para aspecto macroscópico em peças, clique (1)(2)(3)(4)(5)(6). O núcleo é muito bem delimitado da substância branca temporal ao redor. O epêndima do ventrículo lateral aparece logo abaixo. À esquerda na foto, uma pequena veia com espaço perivascular de Virchow-Robin dilatado por artefato de retração. Para o cauda do núcleo caudado em outras técnicas, clique. |
 |
 |
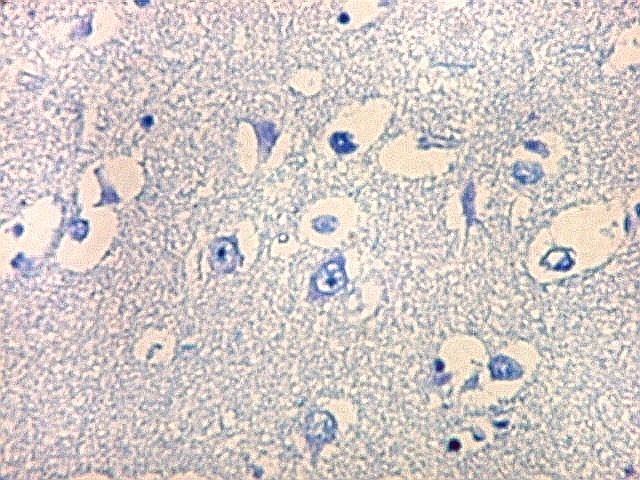
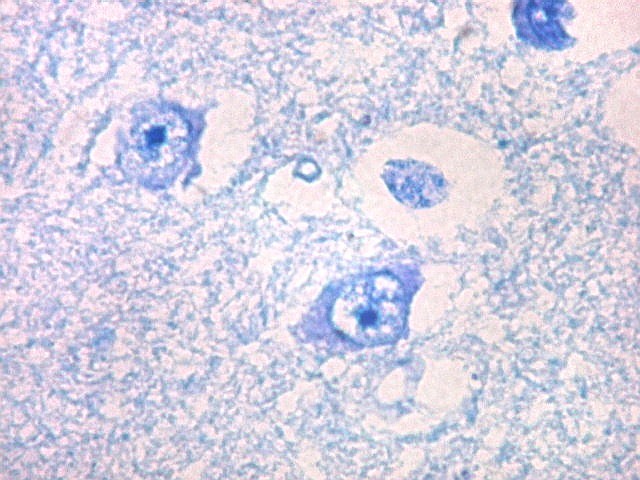
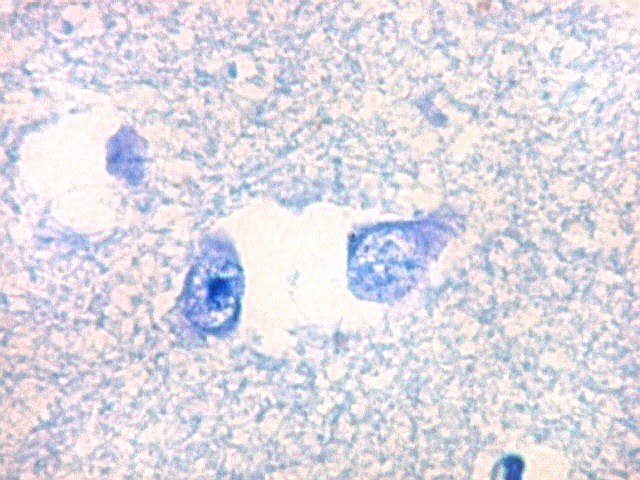
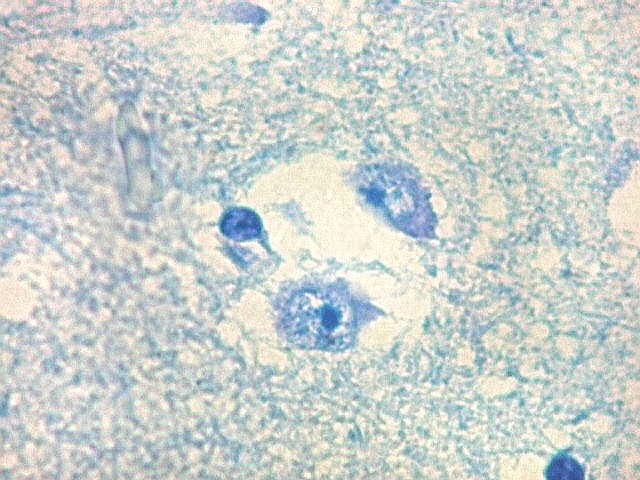
| Núcleo caudado - neurônios. A arquitetura é a mesma de outras regiões do núcleo, inclusive a cabeça e o corpo, situados no centro do encéfalo, ao nível da porção principal do ventrículo lateral. O núcleo caudado é constituído por neurônios pequenos com citoplasma escasso, mas núcleo relativamente grande com nucléolo proeminente. Os dendritos são curtos e pouco evidentes. Os neurônios estão circundados por abundante neurópilo, o tecido intersticial da substância cinzenta rico em prolongamentos celulares de neurônios, sinapses e células da glia. Para neurópilo do córtex cerebral em microscopia eletrônica, clique. Os espaços claros em volta dos corpos celulares dos neurônios são artefatos de retração. | |
 |
 |
 |
 |
 |
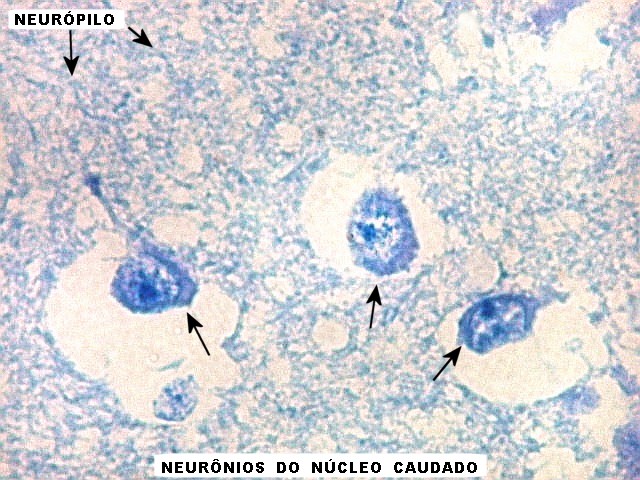 |
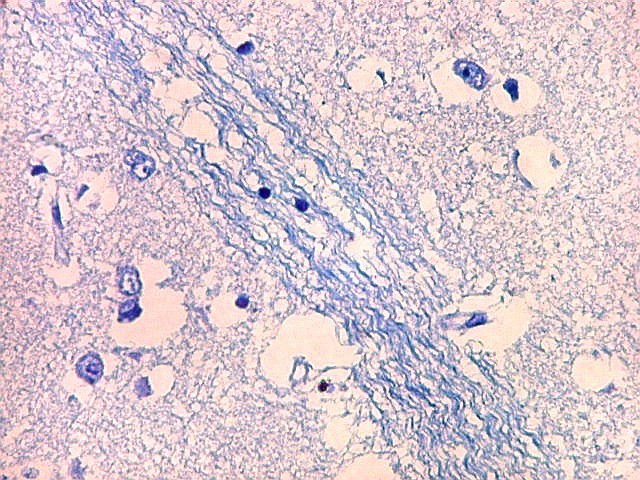
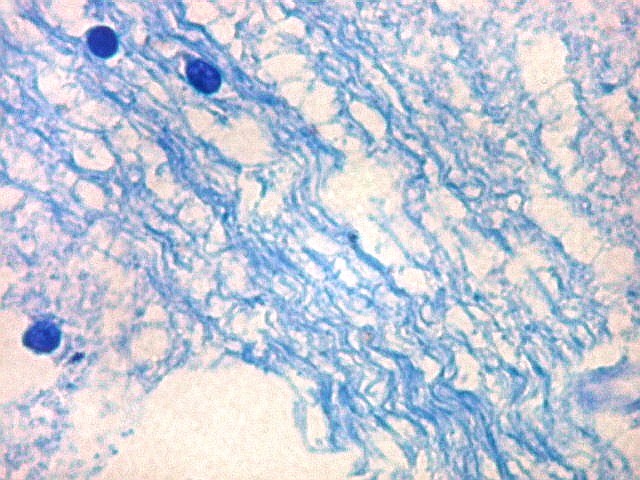
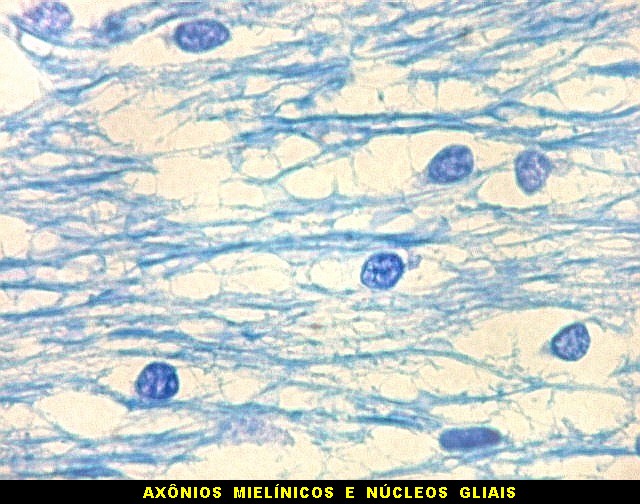
| Núcleo caudado - axônios mielínicos. É característico do núcleo caudado que feixes de axônios mielínicos atravessem a substância cinzenta do núcleo, provenientes da substância branca em torno. Estes se destacam no LFB-Nissl pela estrutura fasciculada e coloração azul turquesa. São extremamente delgados, e entre eles notam-se núcleos da glia, principalmente de oligodendrócitos. |
 |
 |
 |
 |
 |
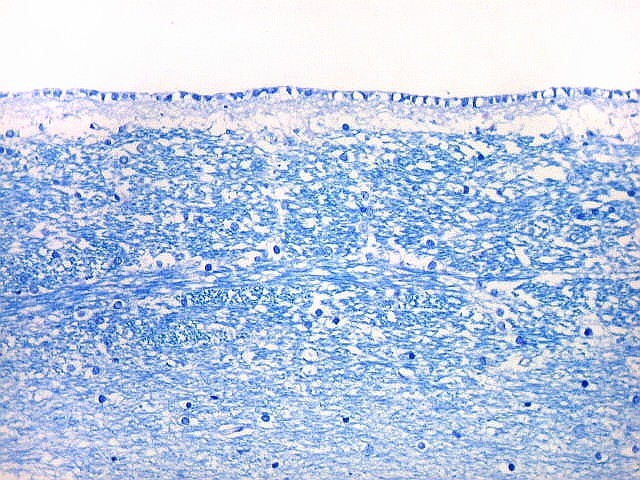
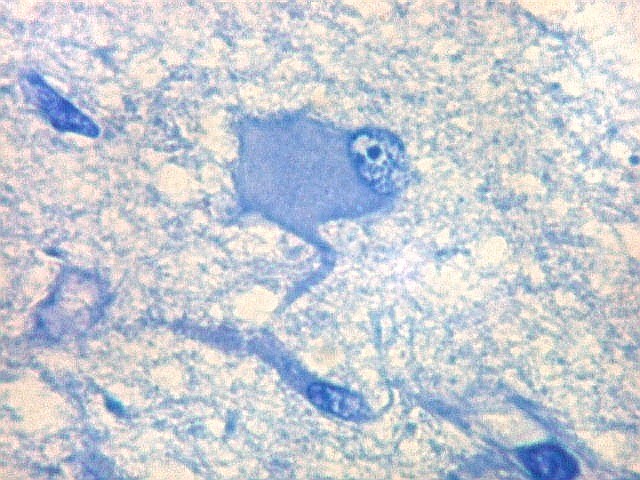
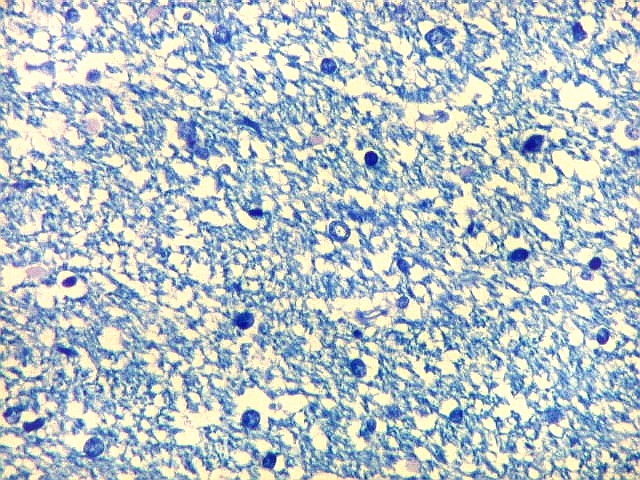
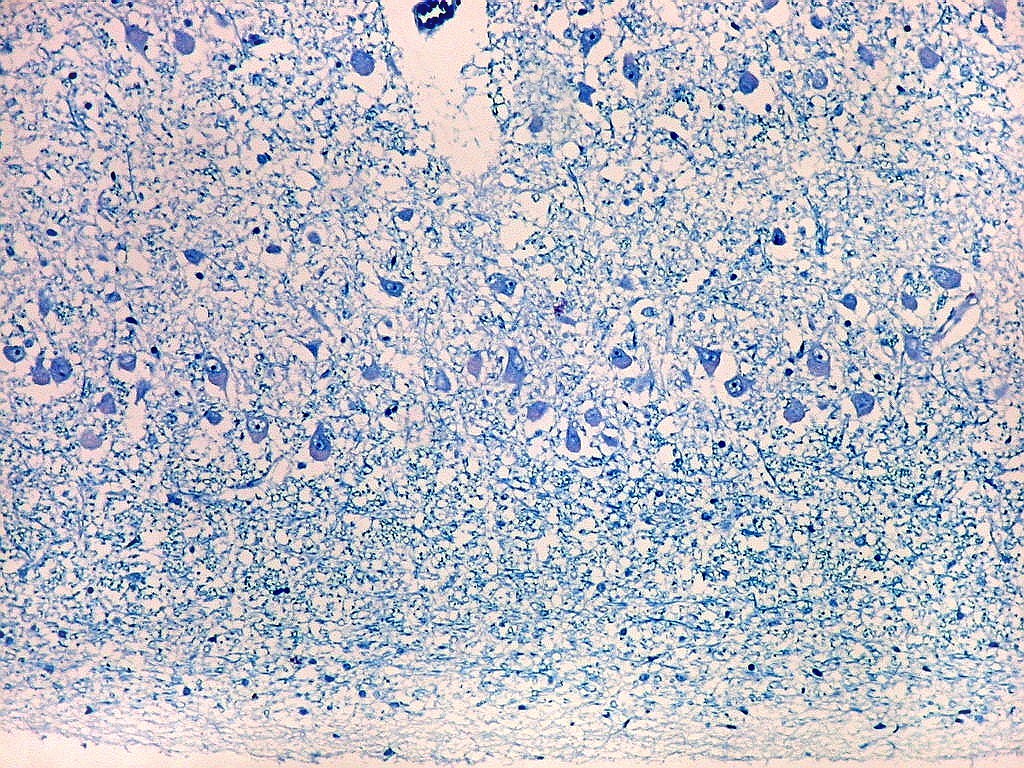
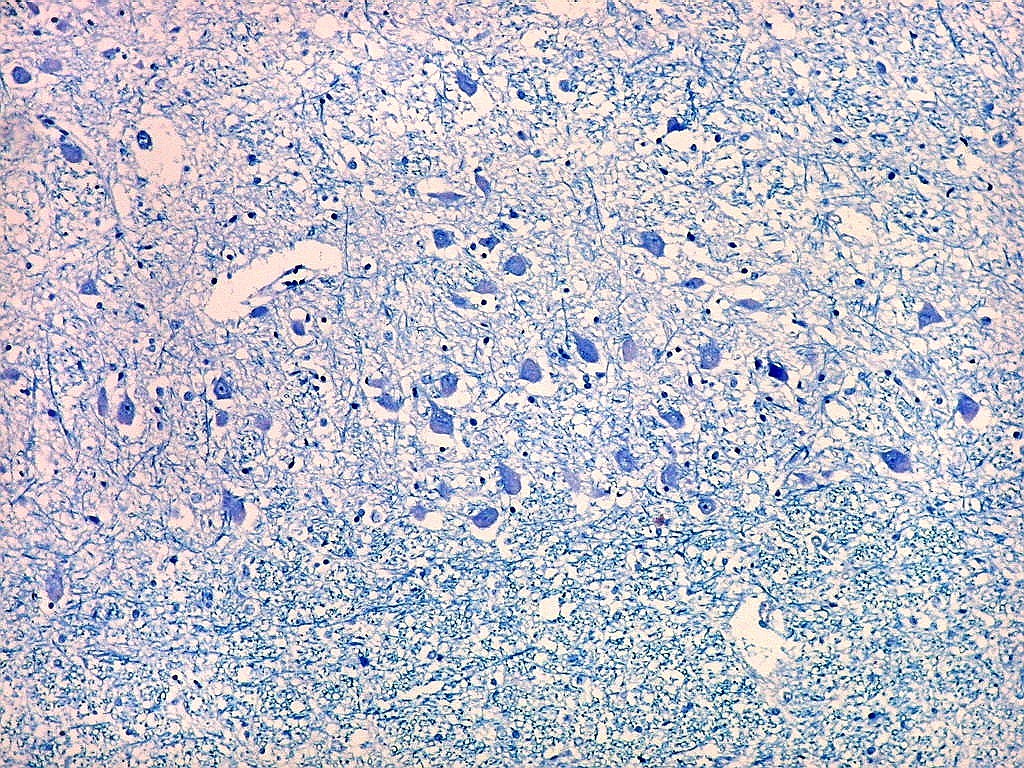
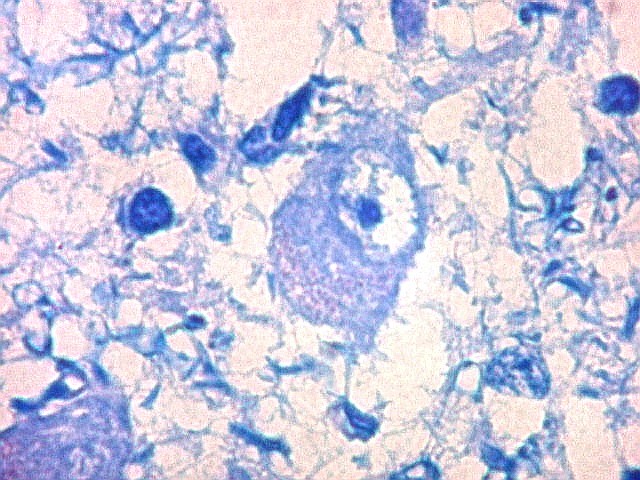
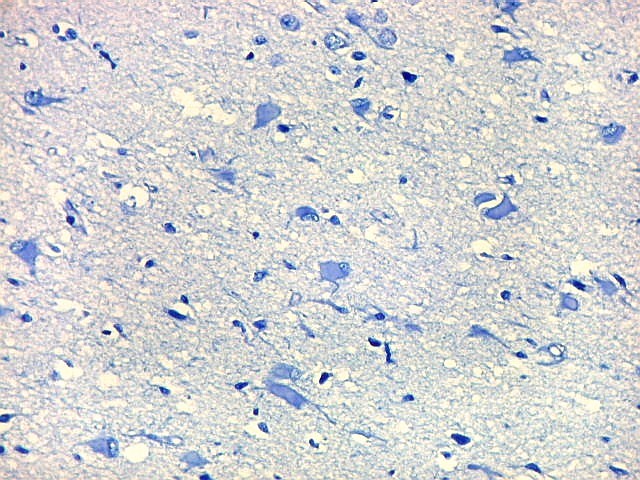
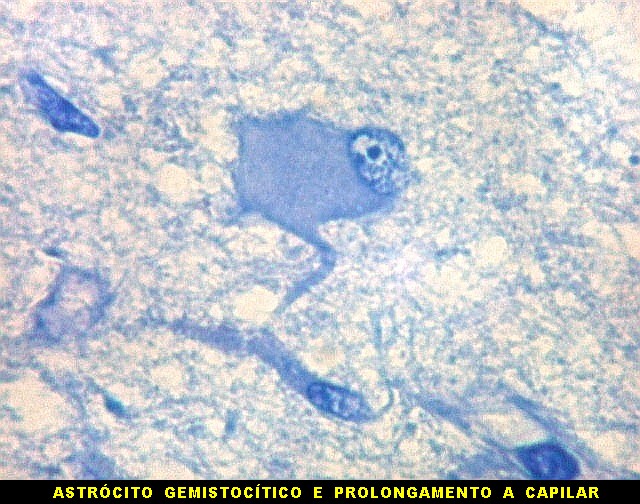
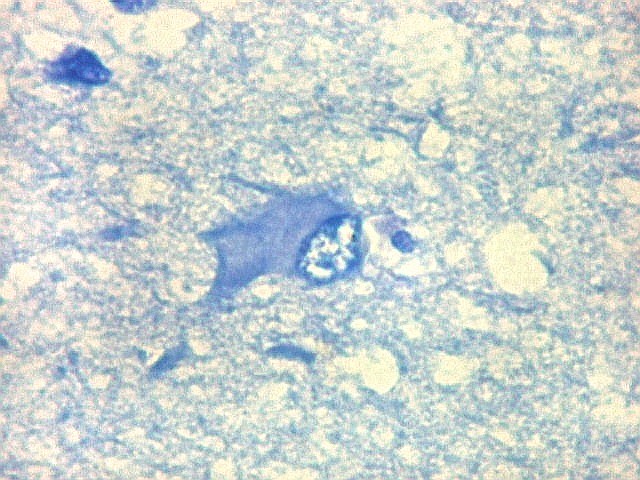
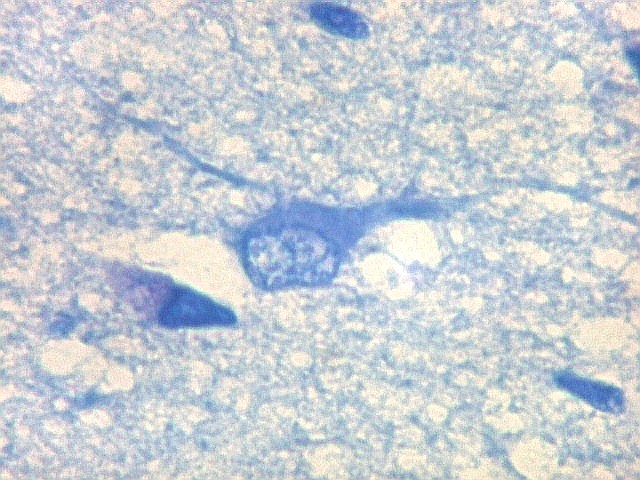
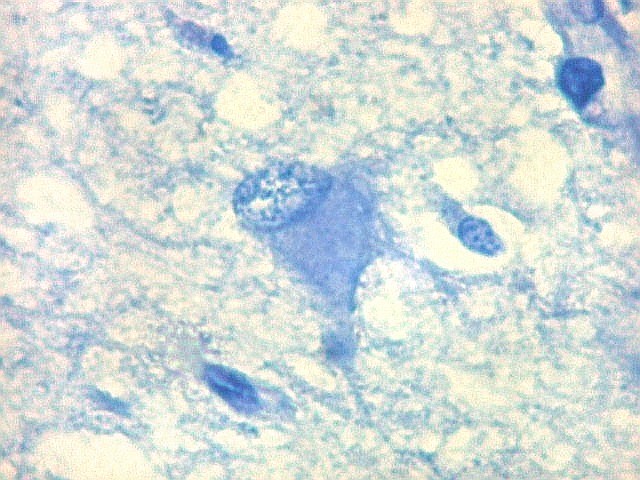
| Área de infarto cortical. Nesta pequena área de infarto no córtex do giro parahipocampal na profundidade do sulco colateral, os neurônios desapareceram. Observam-se astrócitos gemistocíticos, que são formas reacionais a lesões de qualquer tipo no parênquima nervoso. Aparecem como células com citoplasma azul homogêneo e núcleo excêntrico, por vezes com nucléolo proeminente, e prolongamentos que não raro dirigem-se a vasos. Estes astrócitos são bem demonstrados pelas reações imunohistoquímicas para GFAP e VIM. As células microgliais reativas, que aqui pouco aparecem, são proeminentes pelas técnicas para CD68 e HAM-56. | |
 |
 |
 |
 |
 |
 |
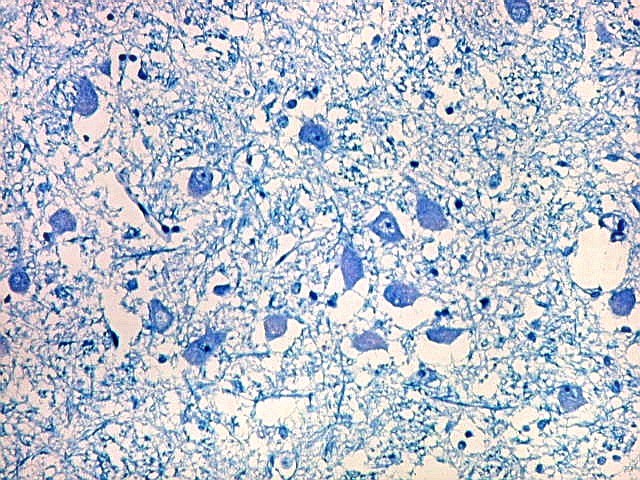
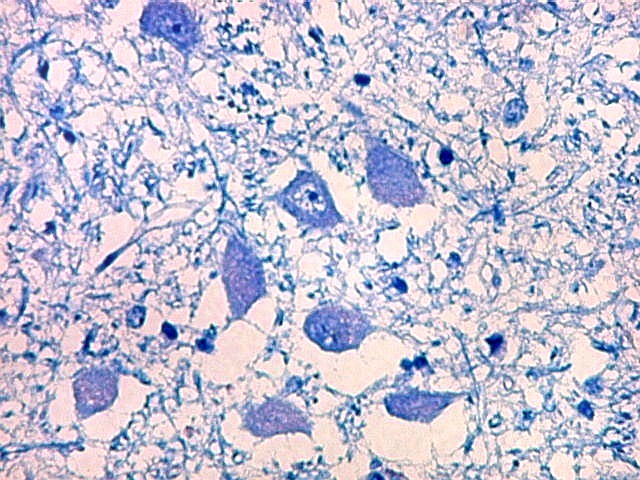
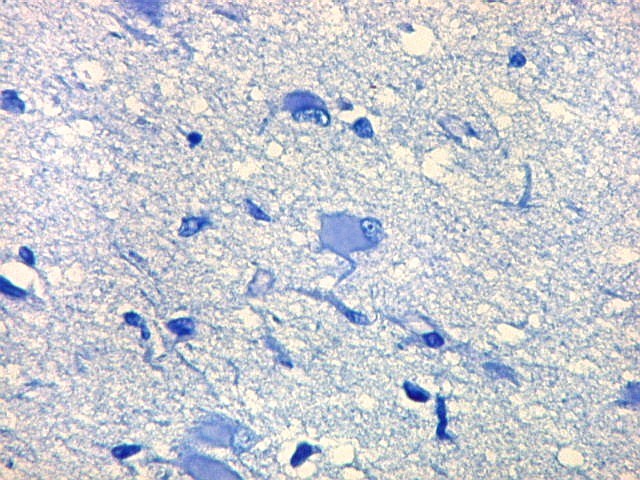
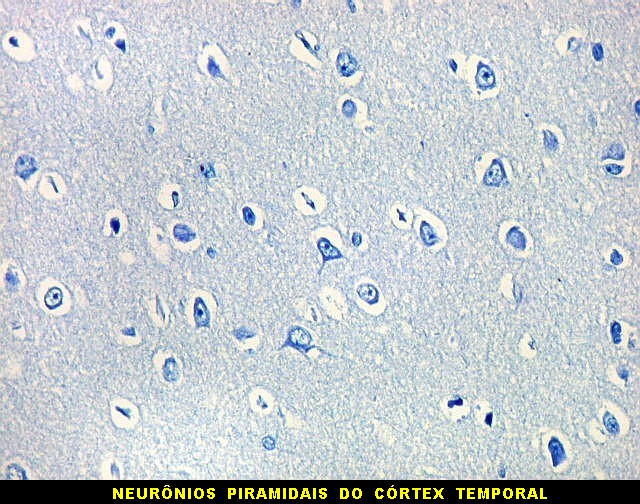
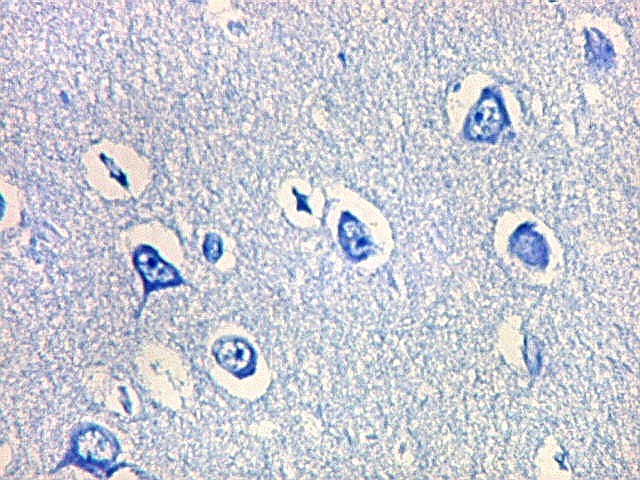
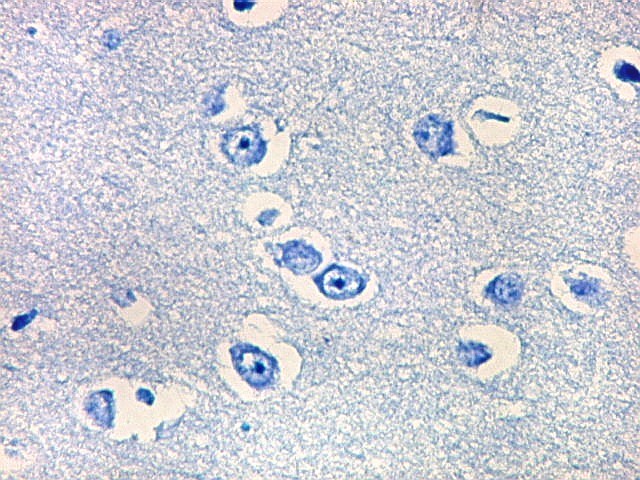
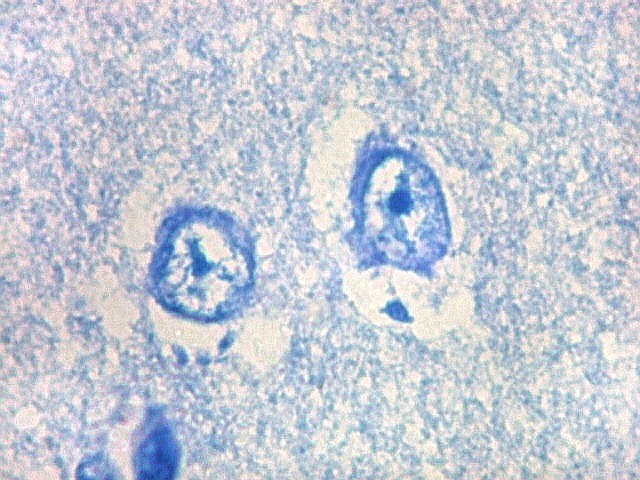
| Neurônios do córtex vizinho. A área de infarto é pequena e bem delimitada e logo faz transição para o córtex normal. Aí observam-se neurônios piramidais clássicos, com formato triangular, núcleo centrado com nucléolo proeminente e citoplasma corado fortemente em azul pelo cresil violeta. Na área infartada, a necrose destes neurônios pela isquemia levou à hipertrofia dos astrócitos e células da micróglia. | |
 |
 |
 |
 |
 |
 |
| Estrutura
do hipocampo.
A formação hipocampal é composta por duas lâminas de córtex de 3 camadas (alocórtex), ambas em forma de U, que se encaixam uma sobre a outra: o hipocampo propriamente dito ou corno de Ammon, e o giro denteado ou fascia dentata. O hipocampo ou corno de Ammon faz proeminência na superfície inferior do corno inferior do ventrículo lateral. O giro denteado fica sob o hipocampo e não aparece externamente, exceto na sua margem medial, chamada margo denticulatus. Na vida embrionária, as duas lâminas (do hipocampo e giro denteado) são uma placa contínua de córtex. A formação do sulco hipocampal na superfície cortical delimita as duas lâminas. A parte profunda do sulco hipocampal logo desaparece devido à fusão das duas lâminas durante a vida fetal. A parte superficial (mais medial) continua visível na superfície medial do lobo temporal. Tanto o hipocampo como o giro denteado são formados do córtex mais simples e primitivo, com 3 camadas (alocórtex ou arquicórtex). A transição para o córtex de 6 camadas ou isocórtex se dá através do peri-alocórtex, formado pelo subículo e zona entorrinal. Estrutura do córtex hipocampal. Da superfície ventricular em direção profunda, isto é, para o sulco hipocampal, o hipocampo apresenta 6 camadas. Excluída a primeira ou alveus, que é substância branca, e condensando-se as 3 últimas em uma, chega-se às 3 camadas clássicas do alocórtex, como discutido abaixo. 1) A camada mais superficial, imediatamente sob o epêndima do corno inferior do ventrículo lateral, é o alveus, composto por uma lâmina de substância branca, que contém os axônios dos neurônios hipocampais e neurônios do subículo. Estes são a principal via eferente de ambas estruturas (hipocampo e subículo) através da fímbria. O alveus também contém fibras aferentes provenientes principalmente da região septal. 2) Segue-se o stratum oriens, que é mal delimitado e mesclado ao stratum pyramidale, que é o seguinte. Contém neurônios em cesto esparsos e os axônios das células piramidais que se dirigem ao alveus. 3) O stratum pyramidale é a principal camada do córtex hipocampal e contém as células piramidais, com corpo tipicamente triangular e base voltada para o alveus. O dendrito apical está voltado para o sulco hipocampal. Os axônios piramidais projetam principalmente para os núcleos septais, mas alguns formam fibras de associação para outros neurônios piramidais e podem cruzar ao hipocampo contralateral. Alguns axônios soltam colaterais de Schaffer, que curvam-se para o stratum radiatum (a seguir) e fazem sinapse nos dendritos apicais de outros neurônios piramidais. Os dendritos apicais dos neurônios piramidais atravessam toda a espessura do córtex hipocampal até o stratum moleculare, junto ao sulco hipocampal. 4) O stratum radiatum consiste dos dendritos apicais dos neurônios piramidais em arranjo paralelo. Estes dendritos são contactados por colaterais de Schaffer, fibras dos núcleos septais e fibras comissurais. 5) O stratum lacunosum contém numerosos fascículos axonais paralelos à superfície do corno de Ammon, formados principalmente por fibras perfurantes e colaterais de Schaffer. 6) O stratum moleculare fica junto ao sulco hipocampal. Devido ao desaparecimento do sulco hipocampal durante o desenvolvimento, o stratum moleculare do corno de Ammon se funde com o do giro denteado. O stratum moleculare tem poucos corpos celulares de neurônios, considerados interneurônios. Contém as arborizações finais dos dendritos apicais dos neurônios piramidais que fazem contato com as colaterais de fibras perfurantes. Como o alocórtex é descrito como de 3 camadas, o alveus é excluído pois é substância branca. As 3 camadas são o stratum oriens, o stratum pyramidale e uma camada formada pela fusão dos strata radiatum, lacunosum e moleculare. Esta camada foi chamada de zona molecular por Ramon y Cajal e Lorente de No. Setores do hipocampo segundo Lorente de No (1934) CA1 a CA4. CA1 é continuação do subículo, sendo os corpos celulares de seus neurônios piramidais tipicamente triangulares, relativamente pequenos e dispersos. CA2 é composto de neurônios grandes, ovóides e densamente agrupados, tornando o stratum pyramidale denso e estreito, em contraste com CA1. Este setor é nítido em hipocampos humanos e de primatas. CA3 corresponde à curva ou joelho do corno de Ammon, onde penetra na concavidade do giro denteado. Os corpos dos neurônios piramidais são semelhantes aos de CA2, mas a densidade é menor. As fibras musgosas do giro denteado circundam os corpos celulares dos neurônios piramidais. Estão também comprimidas entre os estratos radiado e piramidal, formando uma camada suplementar, o stratum lucidum, que é característico de CA3. CA4 está na concavidade do giro denteado. Os corpos neuronais são ovóides, grandes, pouco numerosos e espalhados entre as fibras musgosas e grandes fibras mielínicas características de CA4. Giro denteado. Em cortes transversais pelo corpo do hipocampo, o giro denteado aparece como uma lâmina fina, com concavidade em direção dorsal (ou superior). A concavidade do giro denteado envolve o setor CA4 do hipocampo. O giro denteado é separado dos setores CA1, CA2 e CA3 pelo sulco hipocampal, que desaparece durante o desenvolvimento, embora cavidades residuais possam persistir. Artérias e veias freqüentemente seguem a rota original do sulco hipocampal, separando o corno de Ammon do giro denteado. Na extremidade distal do sulco, as camadas do corno de Ammon e do giro denteado são tão próximas que esta distinção se perde. Uma estreita margem do giro denteado, o margo denticulatus (ou margem denteada) é visível na superfície medial do lobo temporal, sendo o limite do córtex primitivo. O margo denticulatus está separado do subículo pelo sulco hipocampal. O sulco fimbrio-denteado é o limite superficial entre a fímbria e o giro denteado. A estrutura do giro denteado é mais simples que a do corno de Ammon, sendo as 3 camadas do alocórtex facilmente distinguíveis: os estratos molecular e granuloso, e a camada polimórfica. O stratum granulosum, a camada principal, contém os corpos celulares dos neurônios granulosos, que são arredondados, pequenos e densamente agrupados lado a lado. Seus axônios são densamente ramificados, lembrando musgo, daí o nome de fibras musgosas ou mossy fibers, que se dirigem a CA4 e CA3 após atravessar a camada polimórfica. Um dendrito único sai do polo oposto do corpo celular e se estende à camada molecular do giro denteado. Esta é espessa e separada da camada molecular do corno de Ammon pelo sulco hipocampal. Seus dois terços externos próximos ao sulco hipocampal recebem fibras da via aferente perfurante. Seu terço interno, em contato com o estrato granuloso, é ocupado por fibras comissurais e septais. A camada polimórfica ou plexiforme une a camada granulosa do giro denteado a CA4 e é cruzada pelos axônios dos neurônios granulosos. Há poucos neurônios (interneurônios) tanto na camada molecular como na polimórfica. O alocórtex (córtex de 3 camadas) do hipocampo faz transição para o isocórtex do giro parahipocampal através do peri-alocórtex do subículo. O fim do estrato radiado de CA1 é considerado o limite entre o corno de Ammon (hipocampo) e o subículo. O subículo é subdividido em setores. 1 pró-subículo, que continua CA1, e cuja existência não é aceita por todos; 2 subículo propriamente dito, que é parcialmente oculto pelo giro denteado; 3 pré-subículo, que se caracteriza por pequenos neurônios piramidais superficiais agrupados em clusters, que aparecem como pequenas manchas em aumento fraco; 4 parasubículo, contorna a margem do giro parahipocampal para a área entorrinal (área 28 de Brodmann) no aspecto medial do giro. A área entorrinal é mal delimitada. Aceita-se sua presença no uncus e região anterior do giro parahipocampal, mas sua extensão posterior é incerta. São extremamente complexas, ainda mal compreendidas e há numerosas discordâncias entre autores. Aqui, breve sumário baseado na obra de Duvernoy (1988). Vias aferentes principais. São formadas por uma longa cadeia de conexões neuronais, que incluem, sucessivamente, a área entorrinal, o giro denteado, setores CA3 e CA1 do hipocampo e o subículo. A origem está na zona entorrinal, que recebe informações de amplas zonas corticais, particularmente áreas de associação e regiões subcorticais, entre elas tálamo e núcleo amigdalóideo. O claustrum pode ser um intermediário entre o neocórtex e a área entorrinal. A zona entorrinal
manda dois tipos de fibras ao hipocampo: a via alvear e a via perfurante.
As fibras mais numerosas da via perfurante chegam à camada molecular do giro denteado após atravessar o sulco hipocampal. Nos dois terços externos da camada molecular fazem sinapse com dendritos das células granulares. Esta via é a mais aceita por todos autores. Algumas fibras da via perfurante chegam à camada molecular do hipocampo e fazem sinapse nos dendritos apicais dos setores CA1, CA2 e CA3. Esta via não é aceita por todos. A próxima conexão na via aferente é o giro denteado. Os axônios dos neurônios granulosos, chamados fibras musgosas, são glutaminérgicos e contêm zinco em abundância. Atravessam a camada polimórfica e estimulam dendritos de neurônios localizados principalmente em CA3 e também CA4. Os axônios de CA3 e CA4 entram no alveus, e depois na fímbria, a principal via de eferência do hipocampo. Antes, contudo emitem colaterais, ditas colaterais de Schaffer, que atingem os dendritos apicais dos neurônios piramidais de CA1. Desde Ramon y Cajal, os axônios de CA1 têm sido considerados a principal via de saída do hipocampo, via alveus e fímbria. Hoje, admite-se uma via suplementar, pela qual axônios de CA1, via alveus, produzem colaterais que atingem o subículo. Este responderia por fibras glutaminérgicas que formam parte substancial das fibras do alveus e fímbria. A via eferente principal segue pelo fórnix e atinge o núcleo talâmico anterior, diretamente ou através dos corpos mamilares e fascículo mamilo-talâmico. Do tálamo, outras vias atingiriam o hipotálamo e córtex cerebral, particularmente o giro do cíngulo. Por sua vez, o giro do cíngulo projeta diretamente ao hipocampo, além de aferências diretas como dos lobos temporal e pré-frontal. O setor CA2, que é muito proeminente e distinto no hipocampo humano, tem papel obscuro, mas é provável que tenha funções importantes, tendo em vista sua alta celularidade e vascularização. Além das vias principais esquematizadas acima, há circuitos reguladores. Os circuitos reguladores internos têm como elemento básico interneurônios gabaérgicos inibidores, que formam terminações em cesto (basket neurons) em torno dos neurônios piramidais do hipocampo e das células granulares do giro denteado. Estes neurônios em cesto são estimulados por colaterais dos próprios neurônios piramidais e, por sua vez, os inibem. O mesmo ocorre no giro denteado, onde os neurônios em cesto são estimulados por colaterais das fibras musgosas, que são os axônios dos neurônios granulares. Admitem-se ainda vários circuitos envolvendo outros neurotransmissores, que seriam variáveis entre as espécies animais. Há por fim circuitos reguladores externos, envolvendo estruturas extrahipocampais, particularmente os núcleos septais, o hipocampo contralateral via fibras comissurais na comissura anterior e comissura do fórnix. Estas vias comissurais são menos importantes em primatas que em roedores. Terminais de fibras septais são colinérgicas, excitatórias, e atuariam sobre neurônios piramidais e granulosos, diretamente , ou indiretamente através de inibição dos interneurônios. Texto condensado de
|
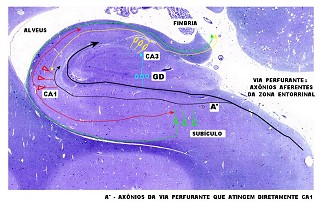
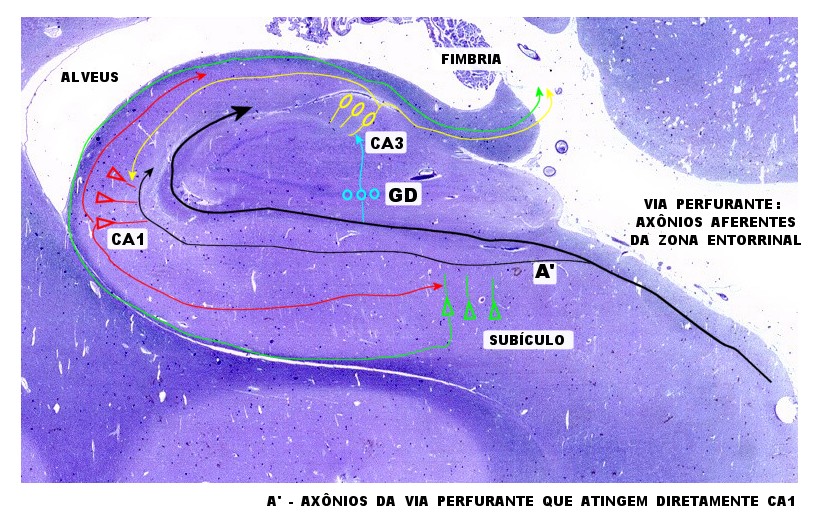
| Esquema
simples das conexões do hipocampo.
Esta versão extremamente simplificada foi redesenhada a partir da
proposta na obra de Duvernoy (1988). Para mais detalhes, clique.
A principal via aferente vem da zona entorrinal, cursa ao longo do sulco hipocampal obliterado, contornando o giro denteado. Quando passam sob o giro denteado (círculos azuis), axônios aferentes fazem sinapse com dendritos de neurônios granulosos. Axônios destes (fibras musgosas) fazem sinapse nos neurônios de CA3 (elipses amarelas). Axônios de CA3 bifurcam, parte entrando no alveus e fímbria. Colaterais destes se dirigem a CA1, onde fazem sinapse nos dendritos dos neurônios piramidais (triângulos vermelhos). Axônios dos neurônios piramidais de CA1 também bifurcam - parte entram no alveus, parte tomam direção oposta para fazer sinapse em neurônios piramidais do subículo (triângulos verdes). Axônios subiculares dirigem-se ao alveus para sair do hipocampo através da fímbria. A via perfurante (A') origina-se de colaterais da via aferente principal que 'perfura' o subículo e cujos axônios fazem contato sináptico diretamente nos dendritos apicais de CA1. |
 |
| Esta simplificação, sem dúvida rudimentar, não leva em conta as conexões com o hipocampo contralateral, vias reguladoras internas e externas, excitatórias e inibitórias. Sua intenção é dar uma visão mínima de uma das estruturas mais complexas e pouco compreendidas do cérebro humano. |
| HE | LFB-Nissl | MAP2 | |
| Página índice da série | 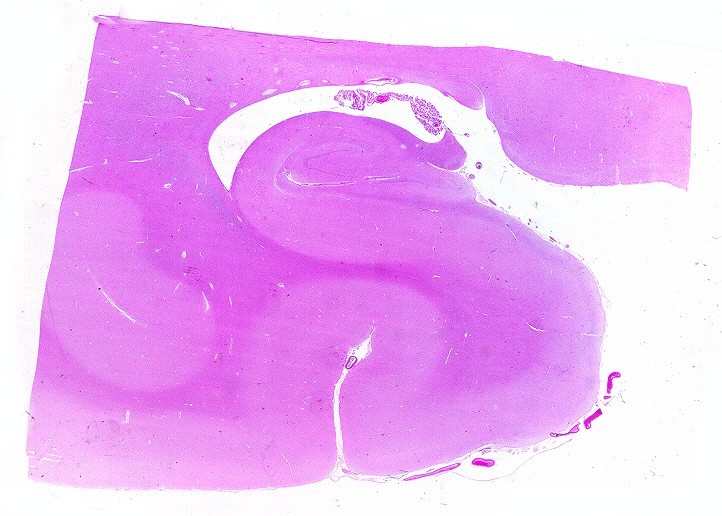 |
|
 |
| NF | SNF | Cromogranina | GFAP |
 |
 |
 |
 |
| VIM | CD34 | CD68 | HAM56 |
 |
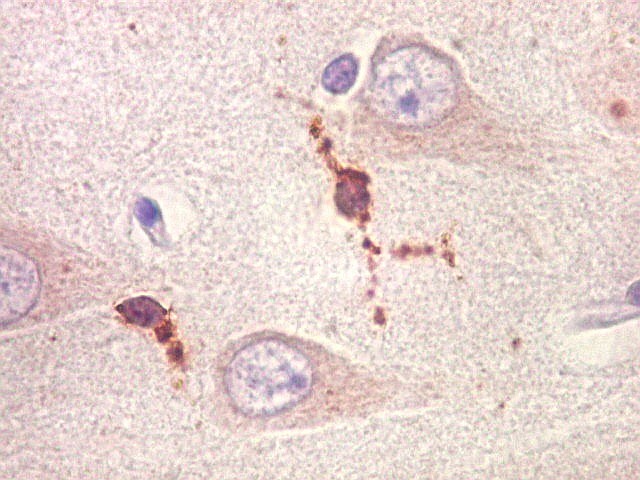
 |
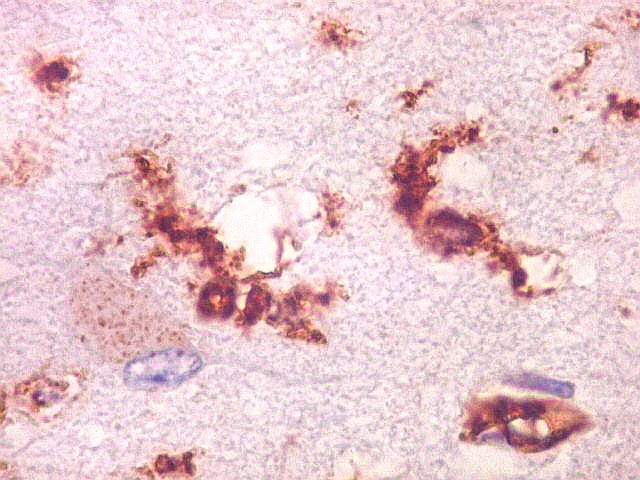
 |
 |
Textos : Estrutura do hipocampo, |
|
||
| Agradecimentos. Cortes histológicos e HE pelo técnico Aparecido Paulo de Moraes. LFB-Nissl pelo técnico Sérgio Roberto Cardoso (in memoriam). Preparações imunohistoquímicas realizadas no Laboratório de Pesquisa pelas técnicas Ana Claudia Sparapani Piaza, Luzia Aparecida Magalhães Ribeiro Reis e Arethusa de Souza. Depto de Anatomia Patológica da FCM-UNICAMP |
| Módulo Neuro - Página Inicial | Outros módulos | e-mails : gradanat@fcm.unicamp.br___gradanat@unicamp.br |