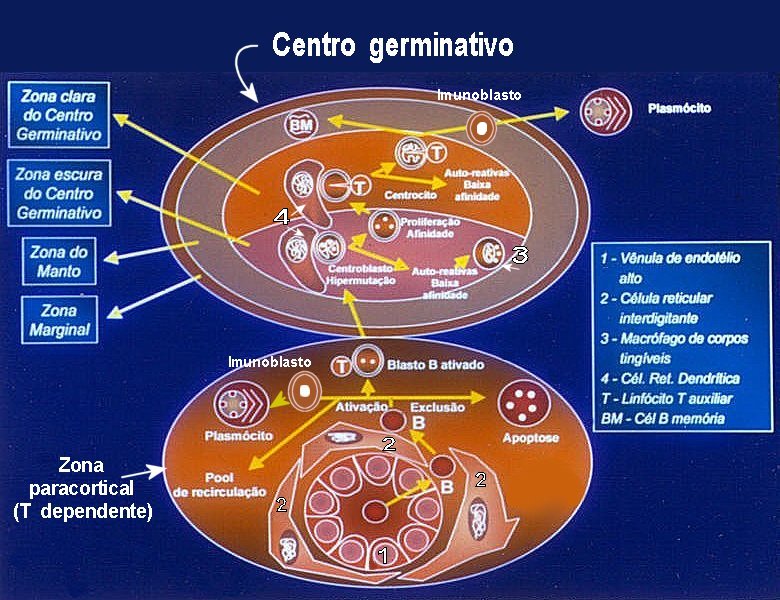
ESQUEMA SIMPLIFICADO
DA PRODUÇÃO DE
CÉLULAS
B EFETORAS E DE MEMÓRIA EM UM
LINFONODO.
No esquema acima há duas elípses. A de baixo representa a
zona
paracortical, T dependente, isto é, a parte da camada
cortical que fica entre os folículos linfóides. A elípse
de cima representa um folículo linfóide, região
B
dependente.
Os linfócitos B ditos virgens, ingênuos ou naïve,
provenientes de células precursoras na medula óssea, vêm
pela circulação sangüínea e, inicialmente, penetram
na zona paracortical ou T através das vênulas de endotélio
alto também ditas vênulas pós-capilares
(1 no esquema). Uma vez atravessada a camada endotelial, têm contato
com as
células reticulares interdigitantes (2 no esquema)
que circundam estas vênulas. Estas células reticulares funcionam
como apresentadoras de antígenos. Ao contactar os antígenos
apresentados, e na dependência do grau de afinidade de seus receptores
de membrana com os ditos antígenos, as células B serão
selecionadas e podem ter quatro destinos:
1) Podem
ser excluídas, sofrendo apoptose. Chama-se isto fenômeno
de exclusão folicular. 2) Podem retornar ao pool de recirculação,
isto é, voltar à corrente sanguínea.
3) Se seus
receptores exibirem boa combinação com o antígeno,
vão sofrer ativação, auxiliadas por uma célula
T. A partir daí sofrem transformação em blastos
B ativados e penetram no centro germinativo, onde ocorrerá
o "refinamento" da célula B. Desta forma serão produzidas
células com capacidade de sintetizar anticorpos com afinidade ainda
maior com o antígeno em questão (ver abaixo).
4) Há
ainda uma quarta possibilidade: se a célula B que entra na zona
paracortical já for uma célula de memória,
esta é ativada ao contato com o antígeno e, mediante atuação
de células T, sofre maturação em plasmócitos.
Neste caso, passa antes pela fase de imunoblasto, uma célula
B imatura cuja característica morfológica é o núcleo
grande com cromatina frouxa e proeminente nucléolo central.
O centro germinativo é polarizado. Tem uma zona mais escura,
voltada para a zona paracortical e para o centro do linfonodo, e uma mais
clara voltada para a periferia. Isto é mais verdadeiro na fase
inicial da ativação do centro germinativo. Posteriormente,
a tendência é de haver só a zona clara. O CG é
circundado por uma zona escura de pequenos linfócitos, chamada zona
do manto e por outra mais fina, a zona marginal. As duas zonas
têm tipos diferentes de células. Ambas são zonas de
passagem, tanto de células que entram (virgens) como de células
que saem (linfócitos B de memória) do centro germinativo,
e ainda contêm células residentes provisórias.
A zona marginal é mais pronunciada na polpa branca do baço
e nos linfonodos mesentéricos. Dela derivam certos linfomas
ditos da zona marginal, como os linfomas MALT e o linfoma esplênico
(ambos indolentes, ver Linfomas).
Ao penetrar inicialmente na zona escura do CG, o blasto B ativado se transforma
em centroblasto, uma célula B cuja característica
morfológica é o núcleo grande com cromatina frouxa
e nucléolos pequenos e múltiplos, geralmente cerca
de três e situados próximos à membrana nuclear.
Os centroblastos estão em contato com as células reticulares
dendríticas do centro germinativo, que são células
apresentadoras de antígenos (número 4 no esquema). Os centroblastos
vão então se multiplicando ativamente por mitose e sofrendo
pequenas mutações que dão às células
filhas a propriedade de produzir anticorpos levemente diferentes dos da
célula mãe. Chama-se a isto hipermutação
somática. Trata-se de mutações pontuais na região
variável dos genes das imunoglobulinas, o que permite gerar a diversidade
dos anticorpos. Os anticorpos são expostos na membrana celular da
célula B, permitindo contato com o antígeno apresentado pelas
célula reticulares dendríticas. Aqueles centroblastos
que produzem anticorpos de pouca afinidade, ou que reagem com antígenos
self
(autoreativos) são excluídos por apoptose. Seus restos são
fagocitados por macrófagos do centro germinativo, ditos macrófagos
de corpos tingíveis (número 3 no esquema), dando
o chamado 'aspecto em céu estrelado'. Os 'corpos tingíveis'
correspondem aos corpos apoptóticos ou restos nucleares das células
que sofreram apoptose.
Por outro lado, os centroblastos que produzem anticorpos de melhor afinidade
serão selecionados para continuar proliferando (seleção
clonal). O processo de refinamento continua na zona clara do CG,
onde os centroblastos se transformam em centrocitos. Estas células
B têm núcleos de cromatina mais condensada e contorno amassado
lembrando uva passa. Não apresentam nucléolos evidentes.
Os centrocitos já estão fora do ciclo celular, isto é,
na fase G0. Contudo podem reativar-se e voltar a se dividir.
Na zona clara há maior concentração de células
T CD4, de grande importantância na seleção clonal.
No fim do processo, a maior parte das células de alta afinidade
com o antígeno apresentarão maturação para
células
B de memória. Estas passam à linfa e retornam à
circulação sanguínea através do ducto torácico.
Uma parcela menor sofrerá transformação em plasmócitos
(células B efetoras), passando pelo estágio intermediário
de imunoblasto, já comentado na zona paracortical. A produção
de plasmócitos é maior na zona medular do linfonodo.
Os imunoblastos migram para os cordões medulares com esta finalidade.